

MEF2 is a converging hub for histone deacetylase 4 and phosphatidylinositol 3-kinase/Akt-induced transformation
- PMID: 24043307
- PMCID: PMC3838174
- DOI: 10.1128/MCB.01050-13
MEF2 is a converging hub for histone deacetylase 4 and phosphatidylinositol 3-kinase/Akt-induced transformation
Abstract
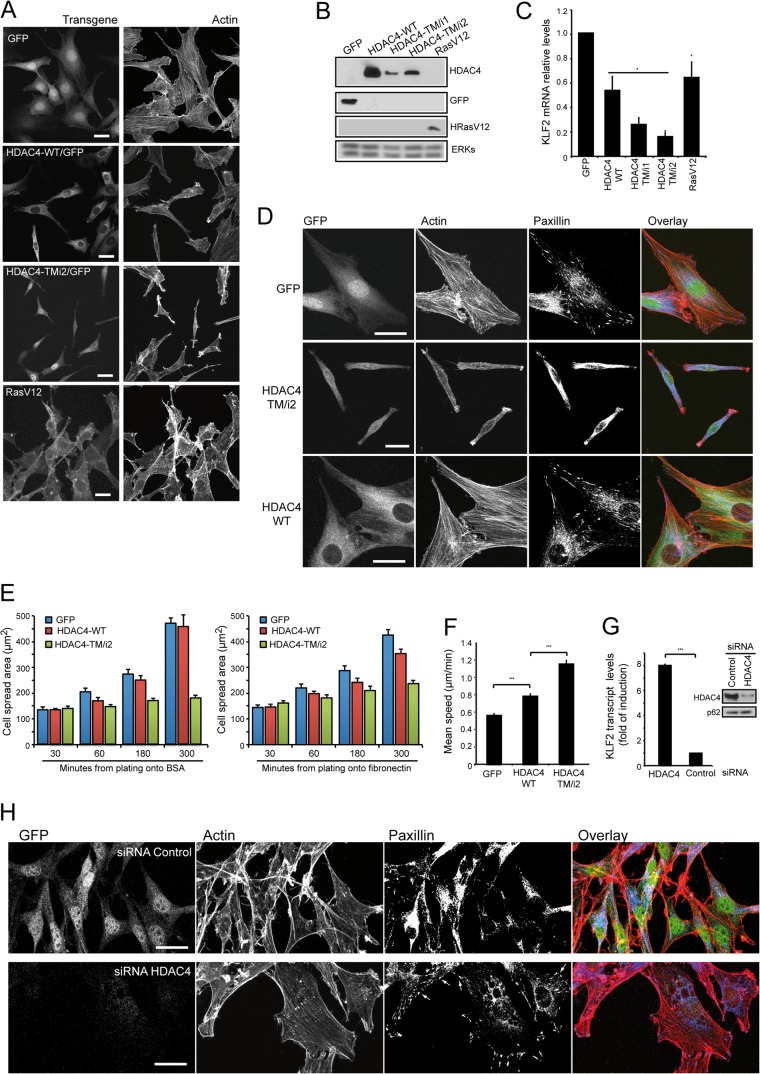
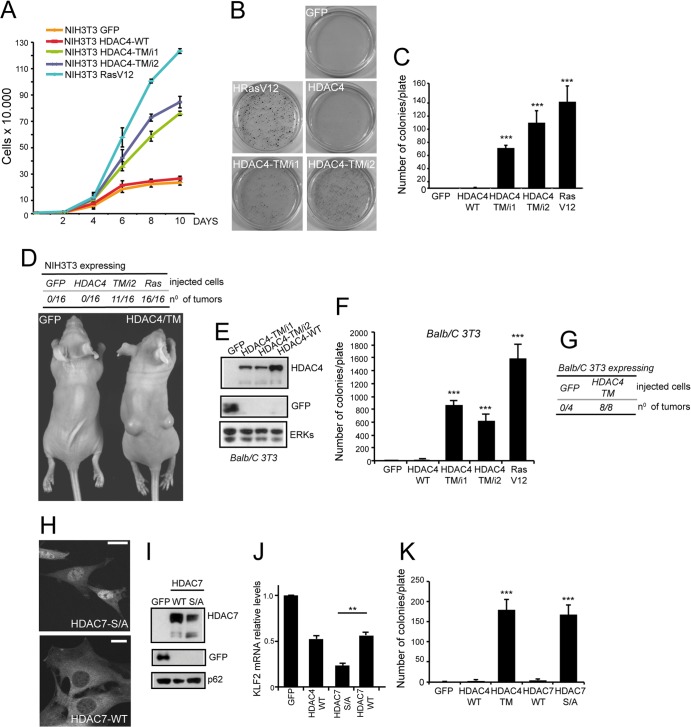
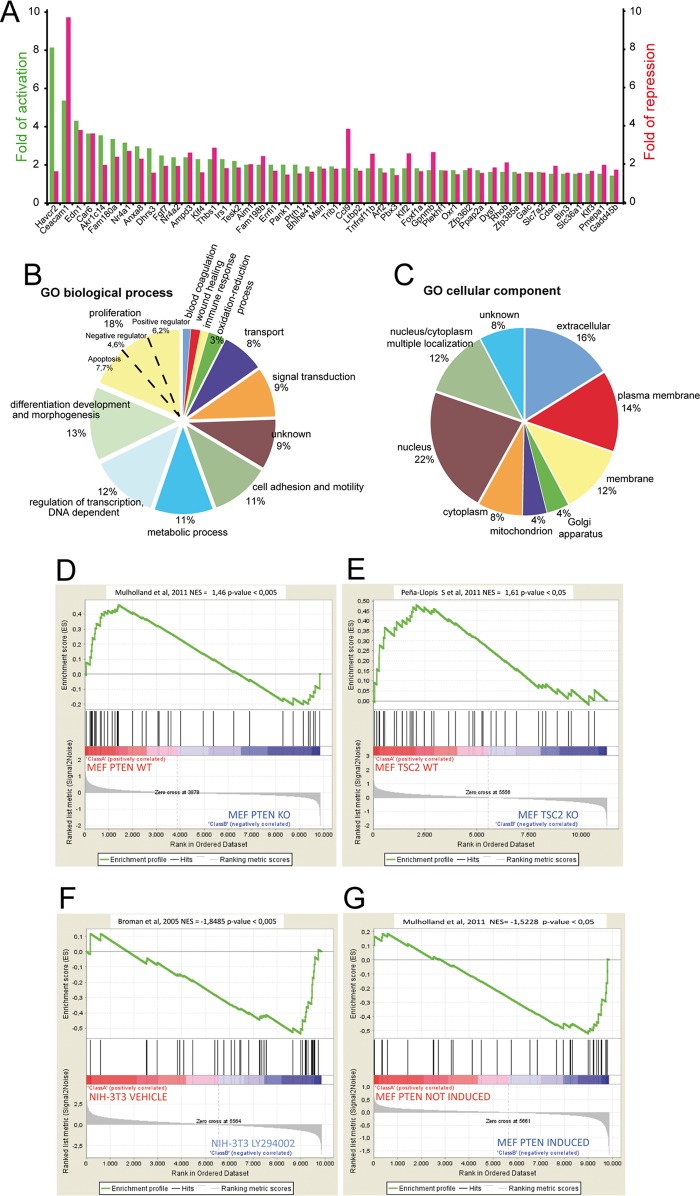
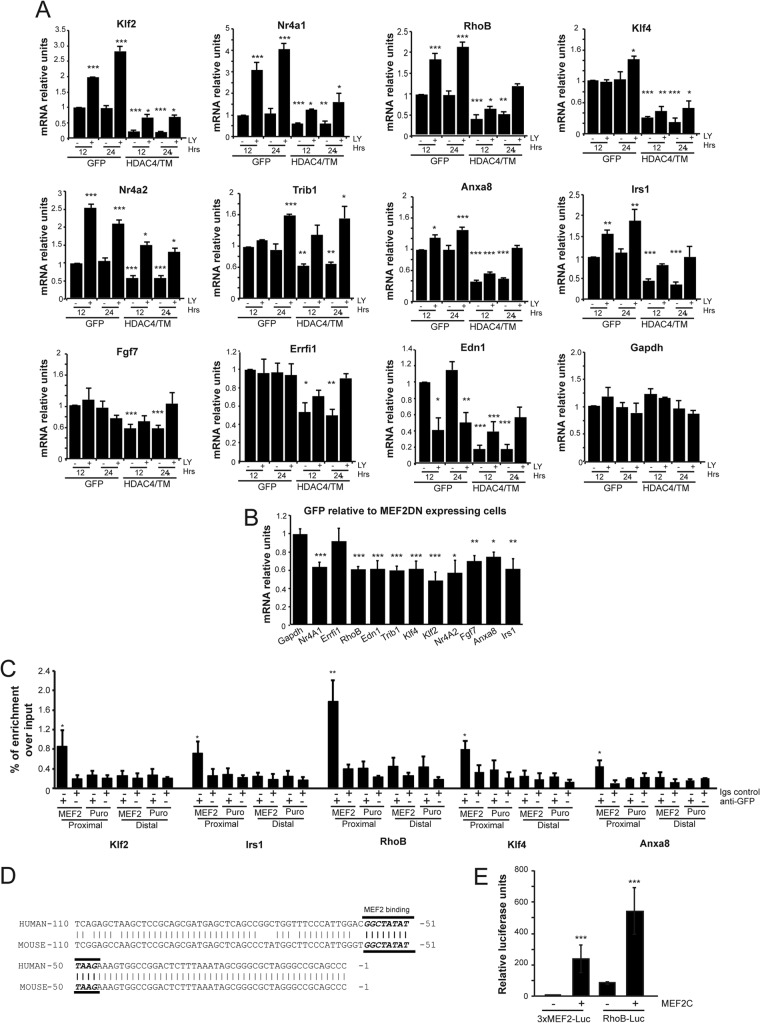
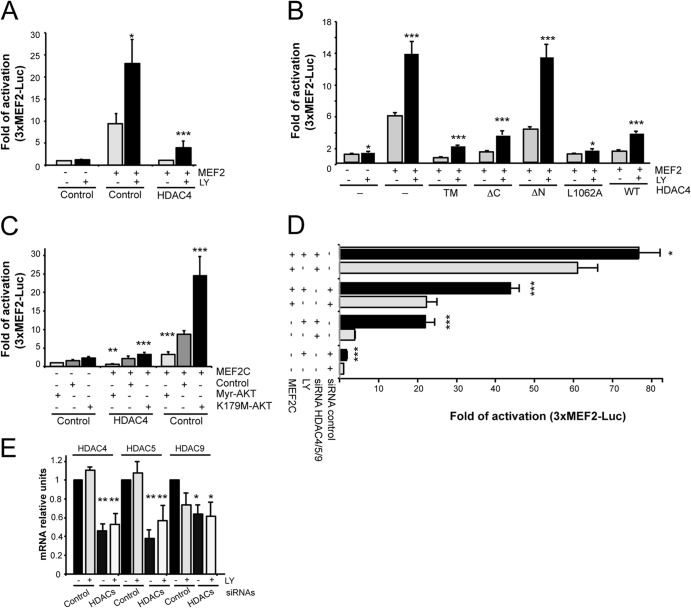
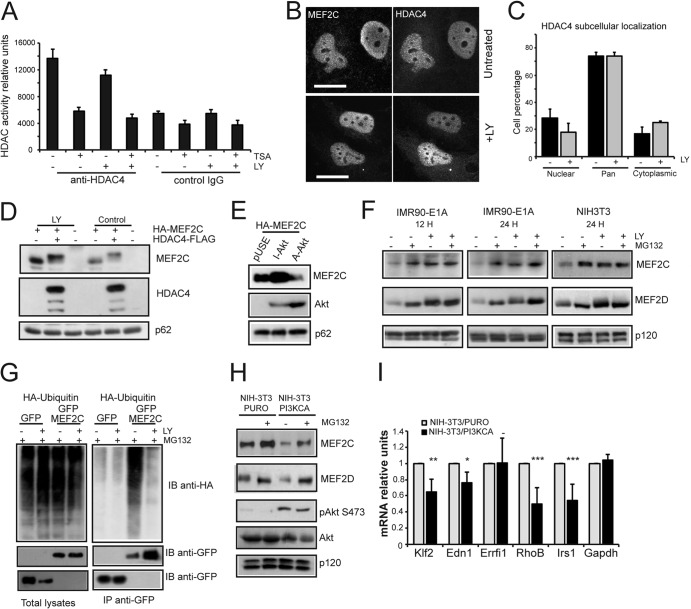
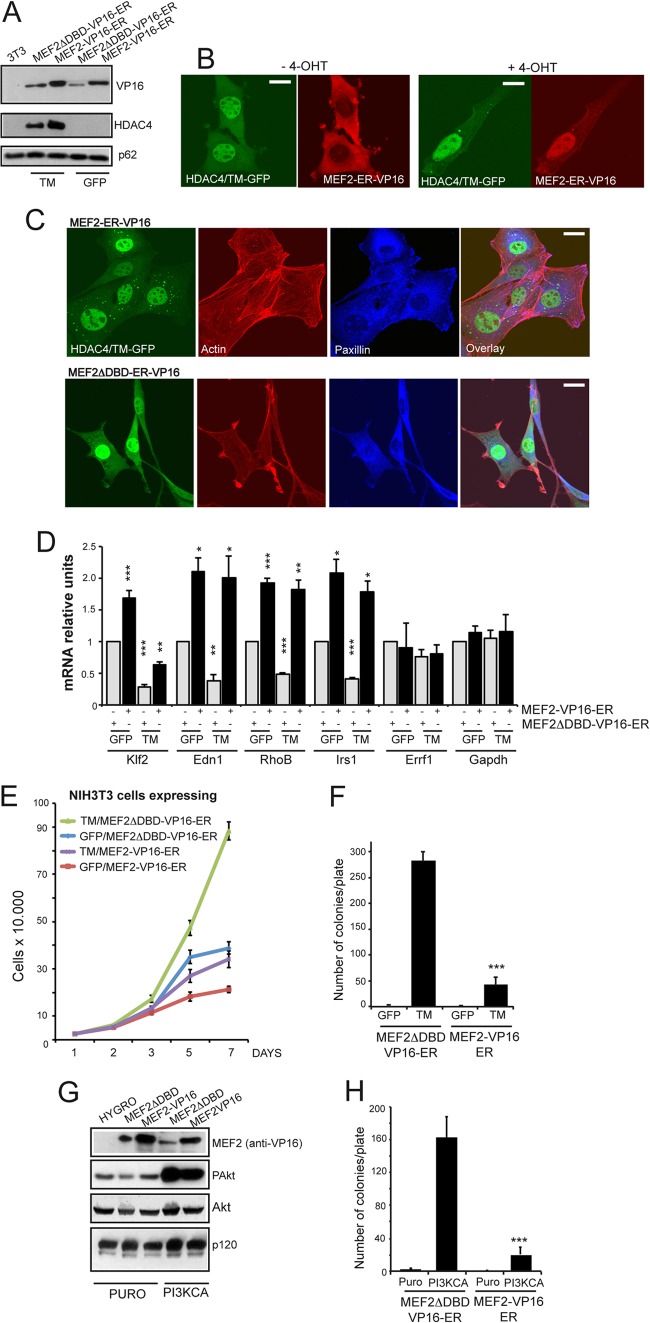
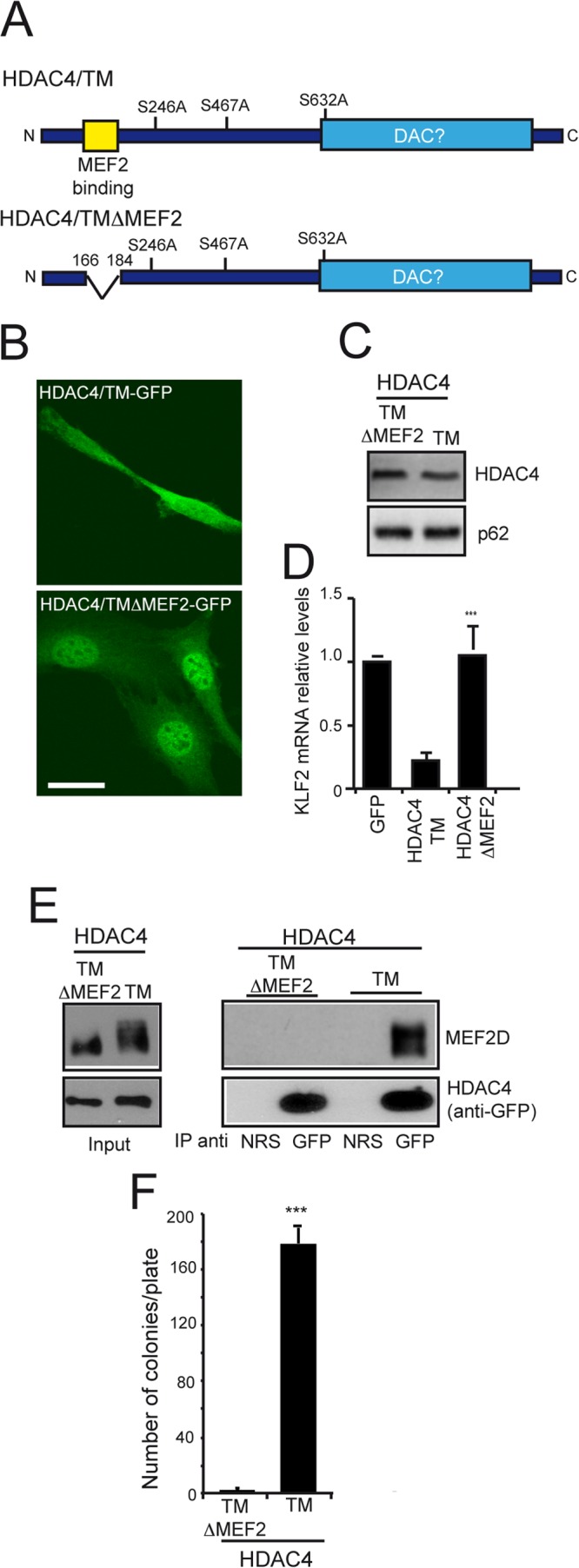
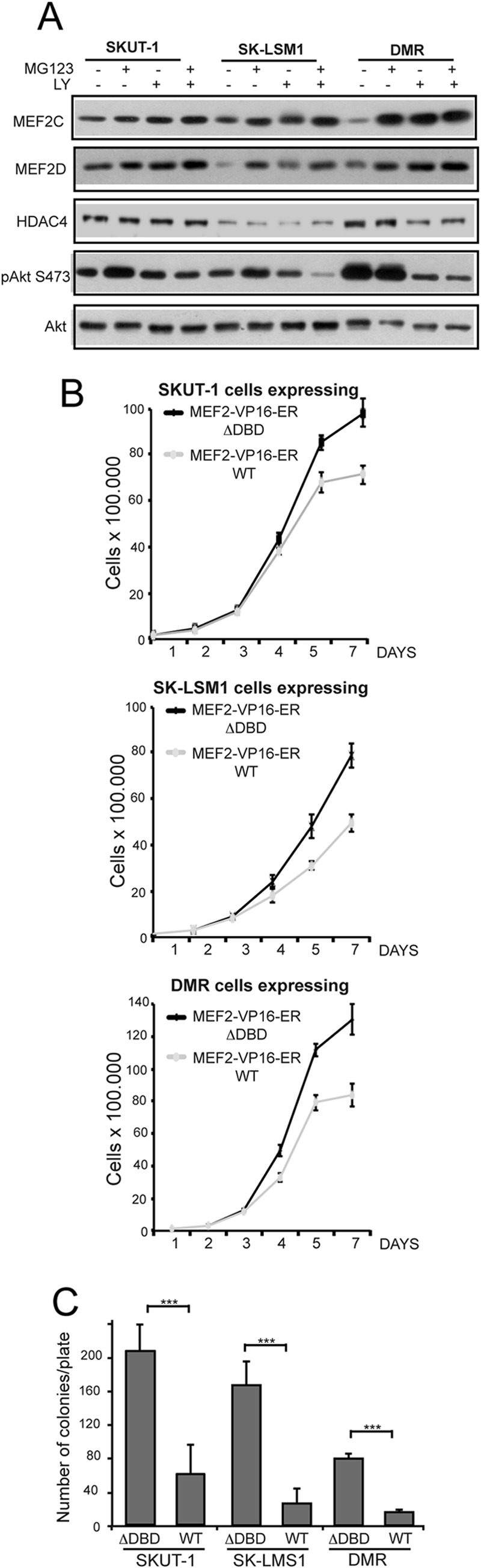
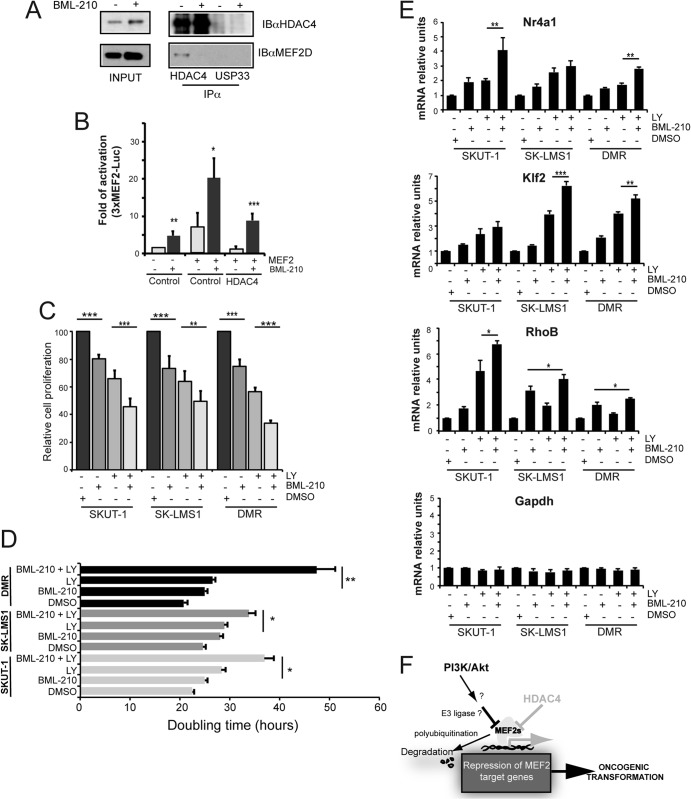
The MEF2-class IIa histone deacetylase (HDAC) axis operates in several differentiation pathways and in numerous adaptive responses. We show here that nuclear active HDAC4 and HDAC7 display transforming capability. HDAC4 oncogenic potential depends on the repression of a limited set of genes, most of which are MEF2 targets. Genes verified as targets of the MEF2-HDAC axis are also under the influence of the phosphatidylinositol 3-kinase (PI3K)/Akt pathway that affects MEF2 protein stability. A signature of MEF2 target genes identified by this study is recurrently repressed in soft tissue sarcomas. Correlation studies depicted two distinct groups of soft tissue sarcomas: one in which MEF2 repression correlates with PTEN downregulation and a second group in which MEF2 repression correlates with HDAC4 levels. Finally, simultaneous pharmacological inhibition of the PI3K/Akt pathway and of MEF2-HDAC interaction shows additive effects on the transcription of MEF2 target genes and on sarcoma cells proliferation. Overall, our work pinpoints an important role of the MEF2-HDAC class IIa axis in tumorigenesis.
Figures

Similar articles
-
The co-existence of transcriptional activator and transcriptional repressor MEF2 complexes influences tumor aggressiveness.PLoS Genet. 2017 Apr 18;13(4):e1006752. doi: 10.1371/journal.pgen.1006752. eCollection 2017 Apr. PLoS Genet. 2017. PMID: 28419090 Free PMC article.
-
Molecular and Functional Characterization of Histone Deacetylase 4 (HDAC4).Methods Mol Biol. 2016;1436:31-45. doi: 10.1007/978-1-4939-3667-0_4. Methods Mol Biol. 2016. PMID: 27246207
-
A dynamic role for HDAC7 in MEF2-mediated muscle differentiation.J Biol Chem. 2001 May 18;276(20):17007-13. doi: 10.1074/jbc.M101508200. Epub 2001 Mar 8. J Biol Chem. 2001. PMID: 11279209
-
Histone deacetylase (HDAC) 9: versatile biological functions and emerging roles in human cancer.Cell Oncol (Dordr). 2021 Oct;44(5):997-1017. doi: 10.1007/s13402-021-00626-9. Epub 2021 Jul 27. Cell Oncol (Dordr). 2021. PMID: 34318404 Free PMC article. Review.
-
The Class IIa histone deacetylase HDAC4 and neuronal function: Nuclear nuisance and cytoplasmic stalwart?Neurobiol Learn Mem. 2015 Sep;123:149-58. doi: 10.1016/j.nlm.2015.06.006. Epub 2015 Jun 11. Neurobiol Learn Mem. 2015. PMID: 26074448 Review.
Cited by
-
HDAC4 degradation during senescence unleashes an epigenetic program driven by AP-1/p300 at selected enhancers and super-enhancers.Genome Biol. 2021 May 10;22(1):129. doi: 10.1186/s13059-021-02340-z. Genome Biol. 2021. PMID: 33966634 Free PMC article.
-
The co-existence of transcriptional activator and transcriptional repressor MEF2 complexes influences tumor aggressiveness.PLoS Genet. 2017 Apr 18;13(4):e1006752. doi: 10.1371/journal.pgen.1006752. eCollection 2017 Apr. PLoS Genet. 2017. PMID: 28419090 Free PMC article.
-
MEF-2 isoforms' (A-D) roles in development and tumorigenesis.Oncotarget. 2019 Apr 12;10(28):2755-2787. doi: 10.18632/oncotarget.26763. eCollection 2019 Apr 12. Oncotarget. 2019. PMID: 31105874 Free PMC article. Review.
-
Third generation quinoline-3-carboxamide transcriptional disrupter of HDAC4, HIF-1α, and MEF-2 signaling for metastatic castration-resistant prostate cancer.Prostate. 2023 Nov;83(15):1470-1493. doi: 10.1002/pros.24606. Epub 2023 Aug 9. Prostate. 2023. PMID: 37559436 Free PMC article.
-
MEF2 plays a significant role in the tumor inhibitory mechanism of encapsulated RENCA cells via EGF receptor signaling in target tumor cells.BMC Cancer. 2018 Dec 4;18(1):1217. doi: 10.1186/s12885-018-5128-5. BMC Cancer. 2018. PMID: 30514247 Free PMC article.
References
-
- Martin M, Kettmann R, Dequiedt F. 2007. Class IIa histone deacetylases: regulating the regulators. Oncogene 26:5450–5467 - PubMed
-
- Fischle W, Dequiedt F, Hendzel MJ, Guenther MG, Lazar MA, Voelter W, Verdin E. 2002. Enzymatic activity associated with class II HDACs is dependent on a multiprotein complex containing HDAC3 and SMRT/N-CoR. Mol. Cell 9:45–57 - PubMed
-
- Lahm A, Paolini C, Pallaoro M, Nardi MC, Jones P, Neddermann P, Sambucini S, Bottomley MJ, Lo Surdo P, Carfí A, Koch U, De Francesco R, Steinkühler C, Gallinari P. 2007. Unraveling the hidden catalytic activity of vertebrate class IIa histone deacetylases. Proc. Natl. Acad. Sci. U. S. A. 104:17335–17340 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials