

Key features of σS required for specific recognition by Crl, a transcription factor promoting assembly of RNA polymerase holoenzyme
- PMID: 24043782
- PMCID: PMC3791729
- DOI: 10.1073/pnas.1311642110
Key features of σS required for specific recognition by Crl, a transcription factor promoting assembly of RNA polymerase holoenzyme
Abstract
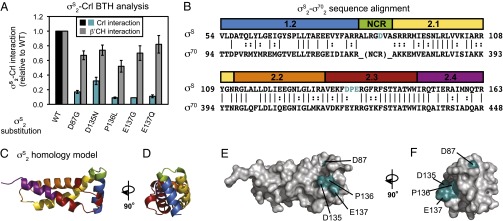
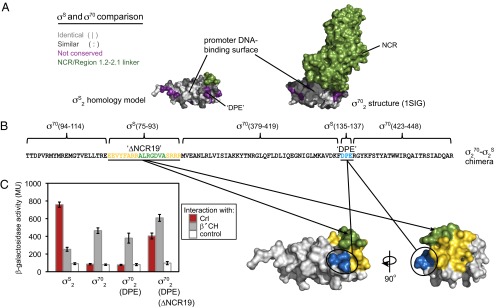
Bacteria use multiple sigma factors to coordinate gene expression in response to environmental perturbations. In Escherichia coli and other γ-proteobacteria, the transcription factor Crl stimulates σ(S)-dependent transcription during times of cellular stress by promoting the association of σ(S) with core RNA polymerase. The molecular basis for specific recognition of σ(S) by Crl, rather than the homologous and more abundant primary sigma factor σ(70), is unknown. Here we use bacterial two-hybrid analysis in vivo and p-benzoyl-phenylalanine cross-linking in vitro to define the features in σ(S) responsible for specific recognition by Crl. We identify residues in σ(S) conserved domain 2 (σ(S)2) that are necessary and sufficient to allow recognition of σ(70) conserved domain 2 by Crl, one near the promoter-melting region and the other at the position where a large nonconserved region interrupts the sequence of σ(70). We then use luminescence resonance energy transfer to demonstrate directly that Crl promotes holoenzyme assembly using these specificity determinants on σ(S). Our results explain how Crl distinguishes between sigma factors that are largely homologous and activates discrete sets of promoters even though it does not bind to promoter DNA.
Keywords: RNAP formation; RpoS; bacterial stress response; curli fiber; transcription initiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Burgess RR, Travers AA, Dunn JJ, Bautz EKF. Factor stimulating transcription by RNA polymerase. Nature. 1969;221(5175):43–46. - PubMed
-
- Gruber TM, Gross CA. Multiple sigma subunits and the partitioning of bacterial transcription space. Annu Rev Microbiol. 2003;57:441–466. - PubMed
-
- Österberg S, del Peso-Santos T, Shingler V. Regulation of alternative sigma factor use. Annu Rev Microbiol. 2011;65:37–55. - PubMed
-
- Lange R, Hengge-Aronis R. Identification of a central regulator of stationary-phase gene expression in Escherichia coli. Mol Microbiol. 1991;5(1):49–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
