

Sensorimotor structure of Drosophila larva phototaxis
- PMID: 24043822
- PMCID: PMC3791751
- DOI: 10.1073/pnas.1215295110
Sensorimotor structure of Drosophila larva phototaxis
Abstract
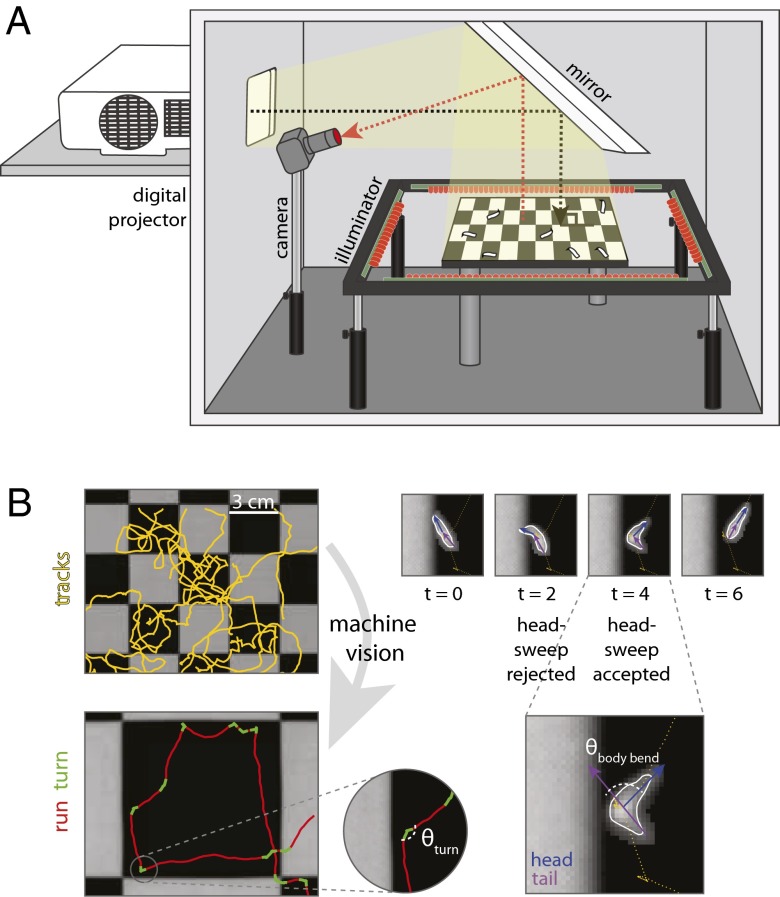
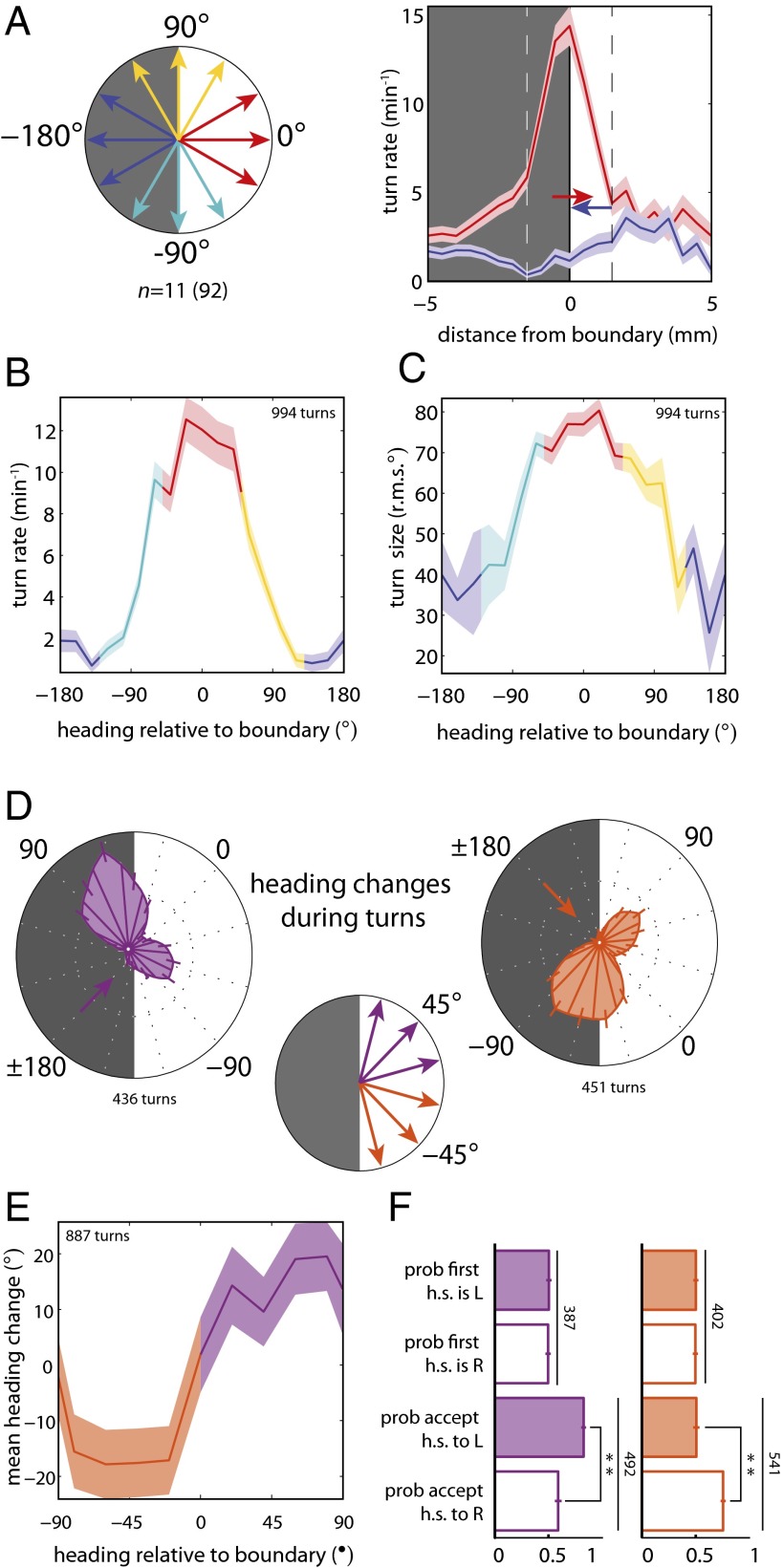
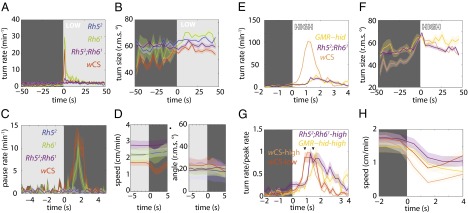
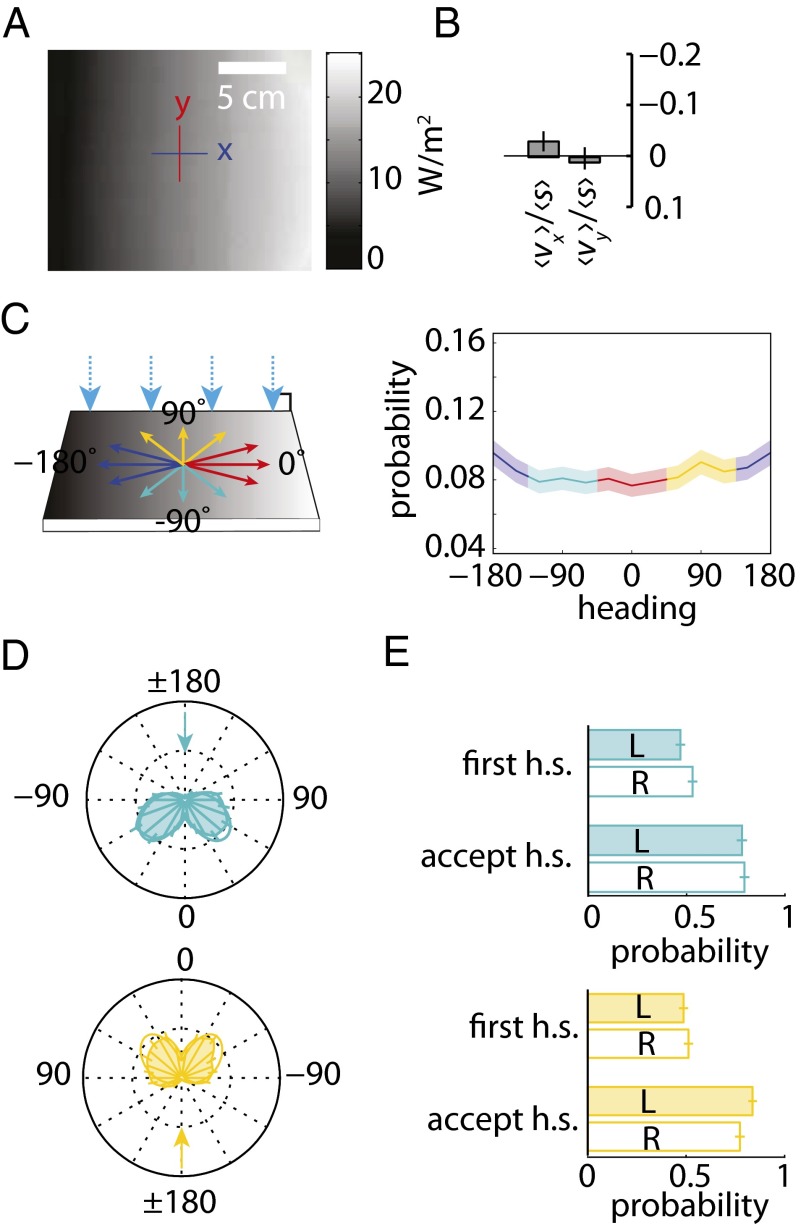
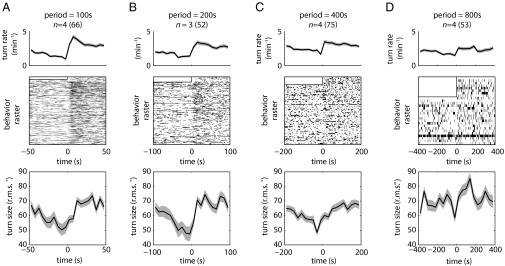
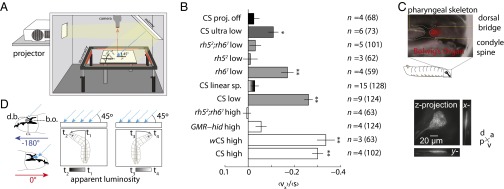
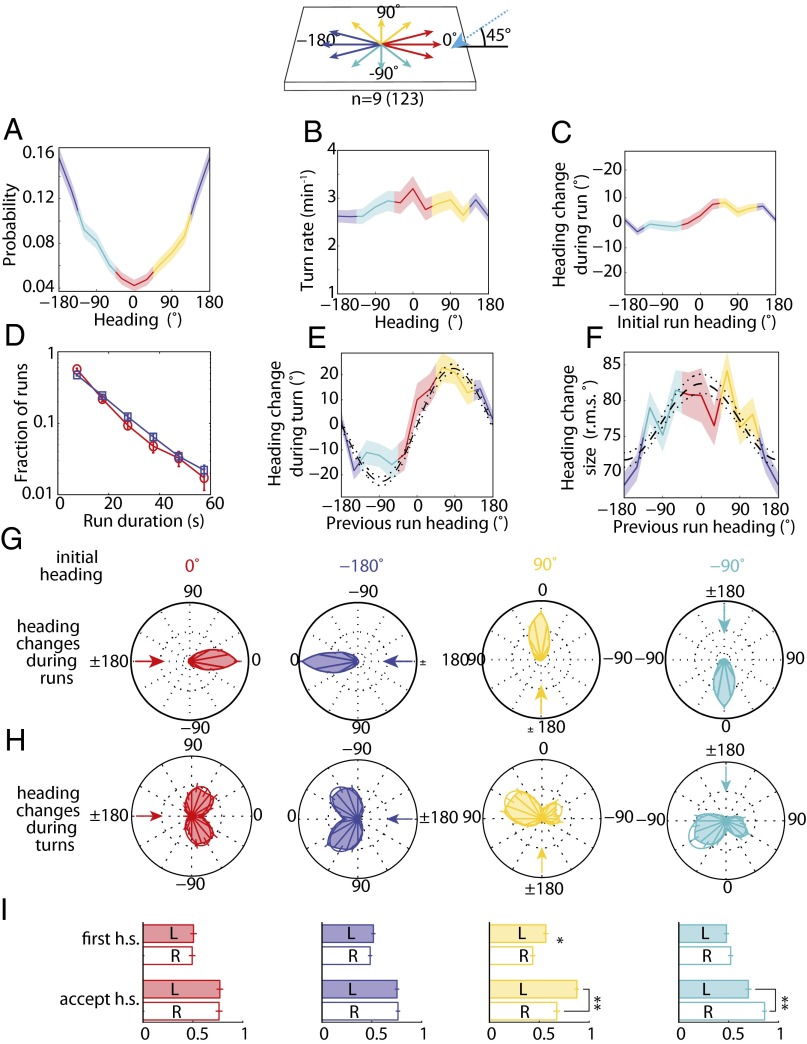
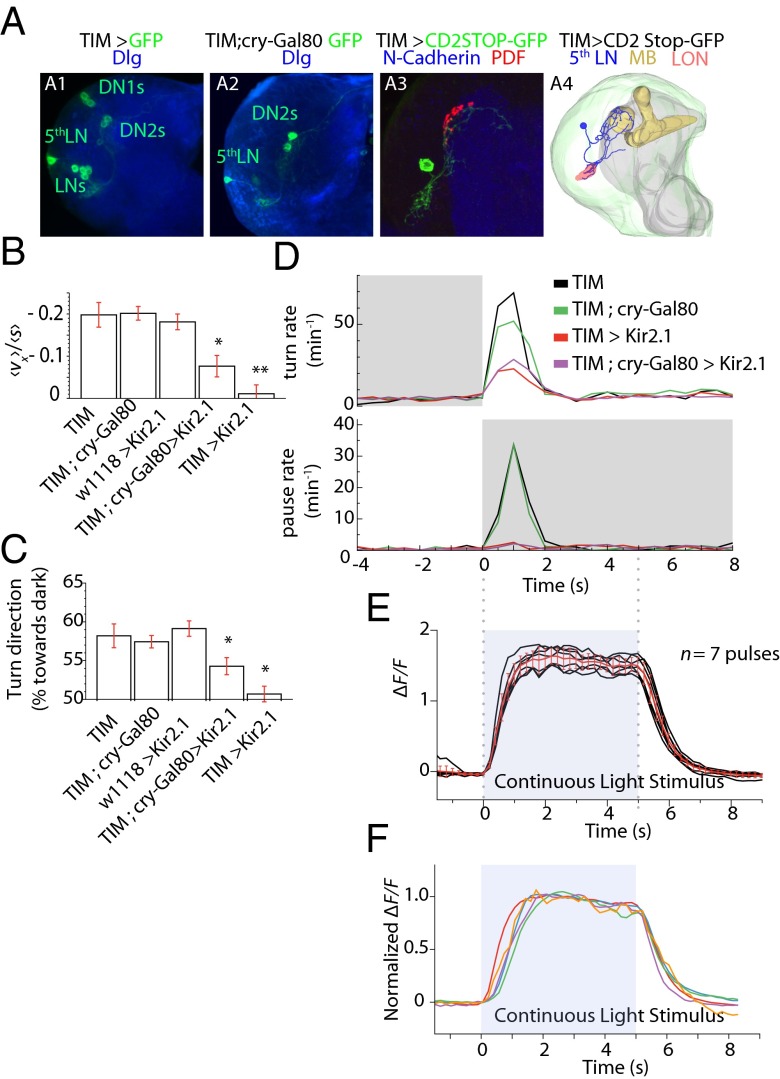
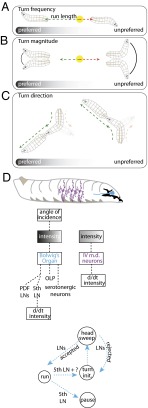
The avoidance of light by fly larvae is a classic paradigm for sensorimotor behavior. Here, we use behavioral assays and video microscopy to quantify the sensorimotor structure of phototaxis using the Drosophila larva. Larval locomotion is composed of sequences of runs (periods of forward movement) that are interrupted by abrupt turns, during which the larva pauses and sweeps its head back and forth, probing local light information to determine the direction of the successive run. All phototactic responses are mediated by the same set of sensorimotor transformations that require temporal processing of sensory inputs. Through functional imaging and genetic inactivation of specific neurons downstream of the sensory periphery, we have begun to map these sensorimotor circuits into the larval central brain. We find that specific sensorimotor pathways that govern distinct light-evoked responses begin to segregate at the first relay after the photosensory neurons.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mast SO. Factors involved in the process of orientation of lower organisms in light. Biol Rev Camb Philos Soc. 1938;13:186–224.
-
- Fraenkel GS, Gunn DL. The Orientation of Animals, Kineses, Taxes and Compass Reactions. Oxford, New York: Oxford University Press; 1961.
-
- Sawin EP, Harris LR, Campos AR, Sokolowski MB. Sensorimotor transformation from light reception to phototactic behavior in Drosophila larvae. J Insect Behav. 1994;7:553–567.
-
- Sawin-McCormack EP, Sokolowski MB, Campos AR. Characterization and genetic analysis of Drosophila melanogaster photobehavior during larval development. J Neurogenet. 1995;10(2):119–135. - PubMed
-
- Hassan J, Busto M, Iyengar B, Campos AR. Behavioral characterization and genetic analysis of the Drosophila melanogaster larval response to light as revealed by a novel individual assay. Behav Genet. 2000;30(1):59–69. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
