

Putative antirecombinase Srs2 DNA helicase promotes noncrossover homologous recombination avoiding loss of heterozygosity
- PMID: 24043837
- PMCID: PMC3791737
- DOI: 10.1073/pnas.1303111110
Putative antirecombinase Srs2 DNA helicase promotes noncrossover homologous recombination avoiding loss of heterozygosity
Abstract
DNA damage alone or DNA replication fork arrest at damaged sites may induce DNA double-strand breaks and initiate homologous recombination. This event can result in a crossover with a homologous chromosome, causing loss of heterozygosity along the chromosome. It is known that Srs2 acts as an antirecombinase at the replication fork: it is recruited by the SUMO (a small ubiquitin-related modifier)-conjugated DNA-polymerase sliding clamp (PCNA) and interferes with Rad51/Rad52-mediated homologous recombination. Here, we report that Srs2 promotes another type of homologous recombination that produces noncrossover products only, in collaboration with PCNA and Rad51. Srs2 proteins lacking the Rad51-binding domain, PCNA-SUMO-binding motifs, or ATP hydrolysis-dependent DNA helicase activity reduce this noncrossover recombination. However, the removal of either the Rad51-binding domain or the PCNA-binding motif strongly increases crossovers. Srs2 gene mutations are epistatic to mutations in the PCNA modification-related genes encoding PCNA, Siz1 (a SUMO ligase) and Rad6 (a ubiquitin-conjugating protein). Knocking out RAD51 blocked this recombination but enhanced nonhomologous end-joining. We hypothesize that, during DNA double-strand break repair, Srs2 mediates collaboration between the Rad51 nucleofilament and PCNA-SUMO and directs the heteroduplex intermediate to DNA synthesis in a moving bubble. This Rad51/Rad52/Srs2/PCNA-mediated noncrossover pathway avoids both interchromosomal crossover and imprecise end-joining, two potential paths leading to loss of heterozygosity, and contributes to genome maintenance and human health.
Keywords: NHEJ; SDSA; bubble migration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
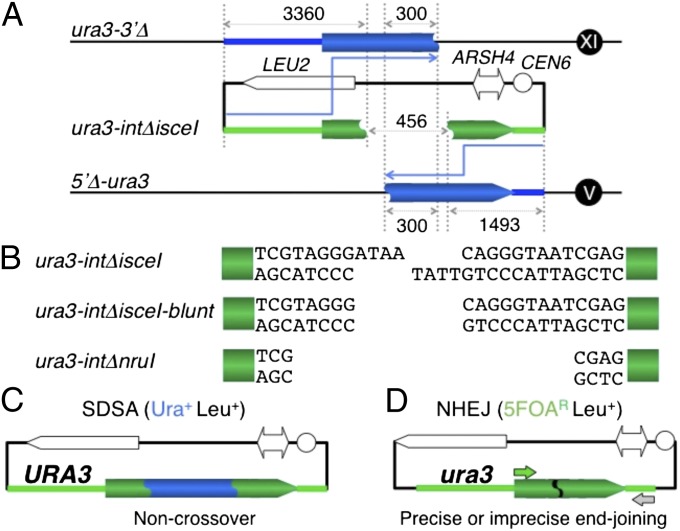
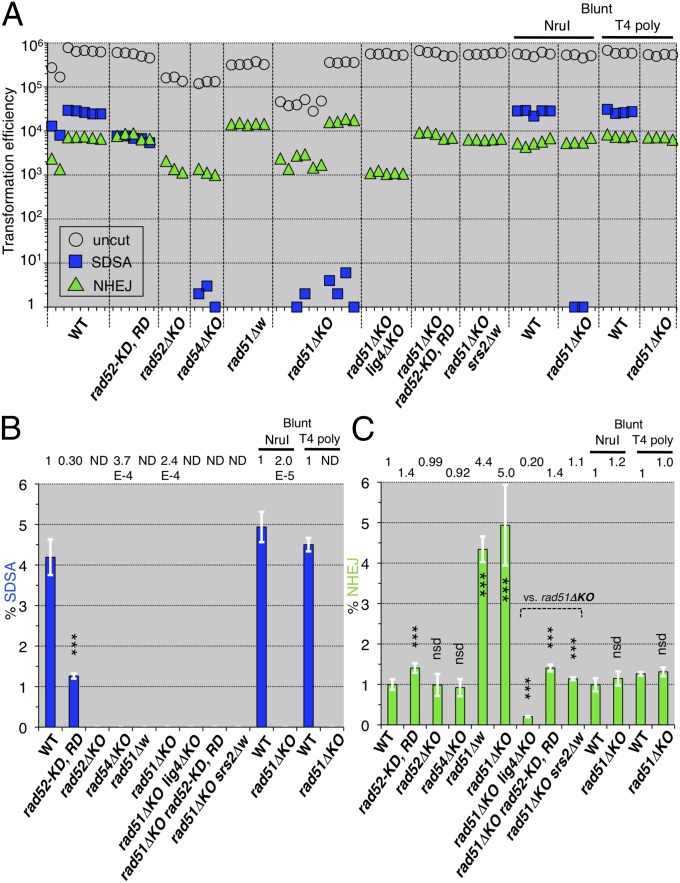
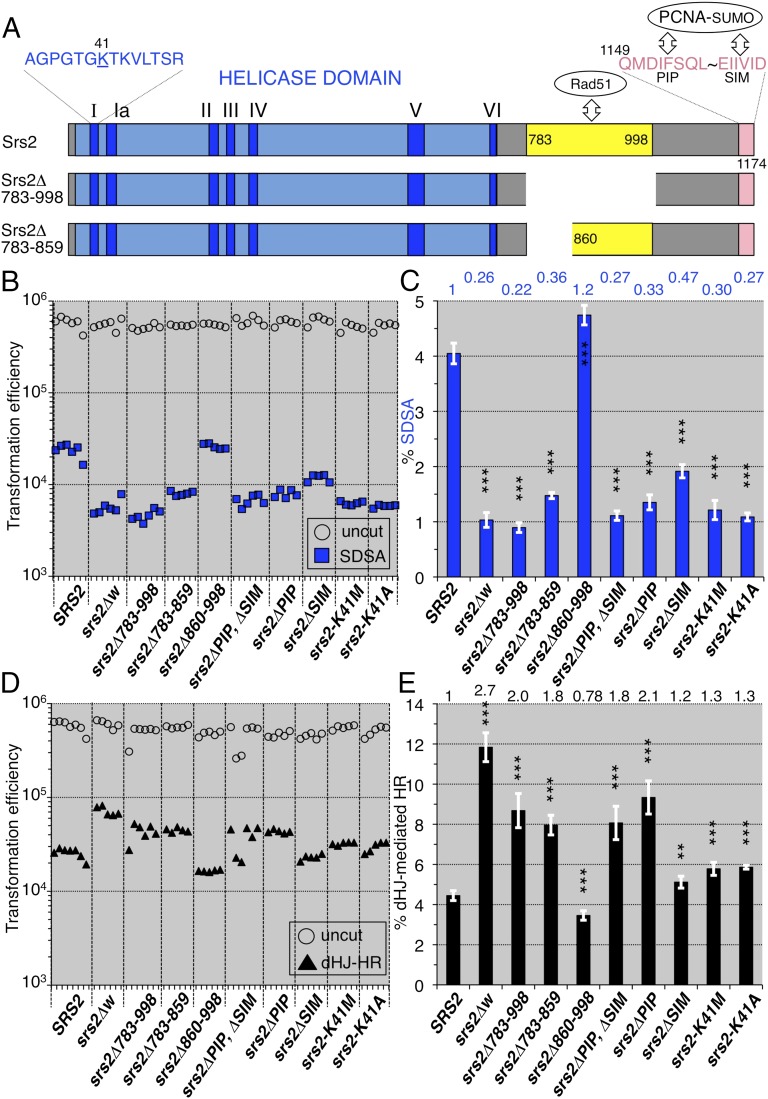
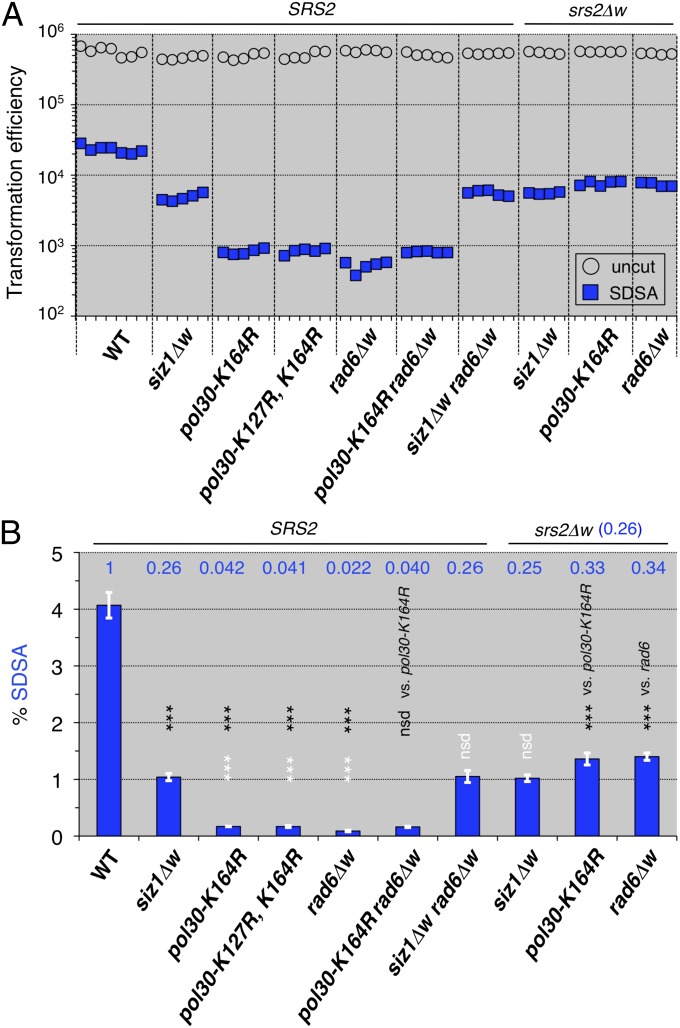
References
-
- Pfander B, Moldovan GL, Sacher M, Hoege C, Jentsch S. SUMO-modified PCNA recruits Srs2 to prevent recombination during S phase. Nature. 2005;436(7049):428–433. - PubMed
-
- Krejci L, et al. DNA helicase Srs2 disrupts the Rad51 presynaptic filament. Nature. 2003;423(6937):305–309. - PubMed
-
- Veaute X, et al. The Srs2 helicase prevents recombination by disrupting Rad51 nucleoprotein filaments. Nature. 2003;423(6937):309–312. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
