

The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis
- PMID: 24045019
- PMCID: PMC3809544
- DOI: 10.1105/tpc.113.117168
The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis
Abstract
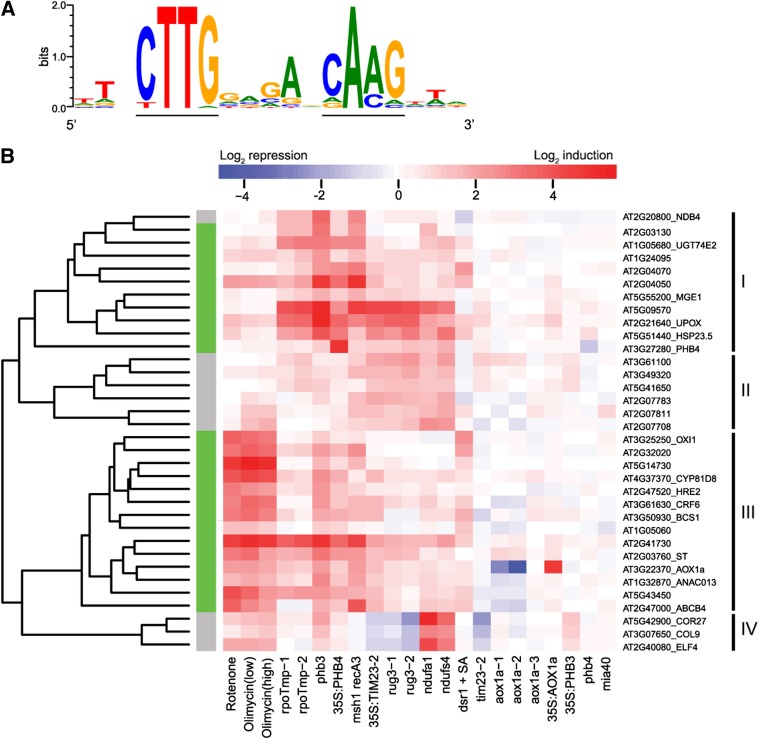
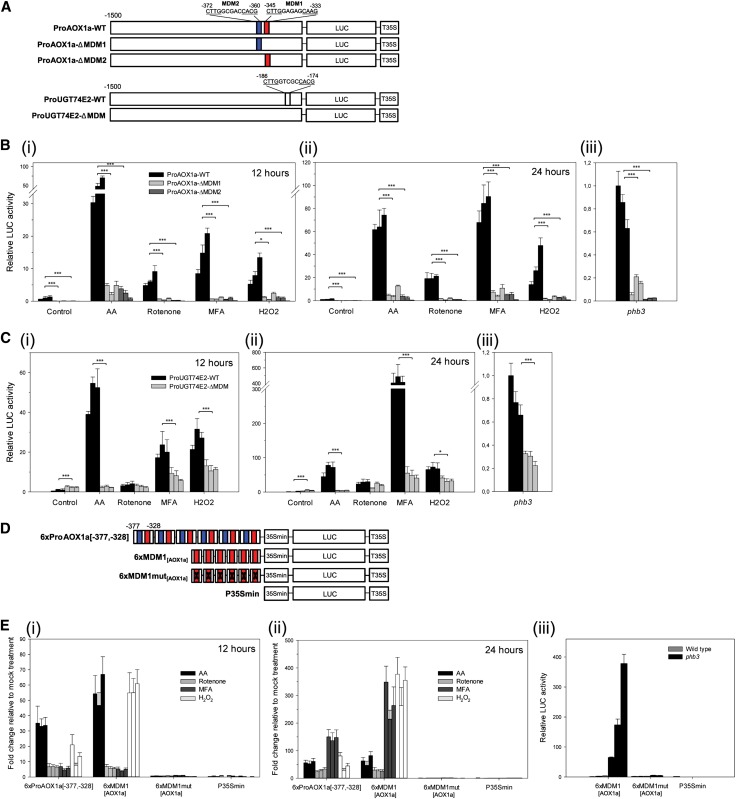
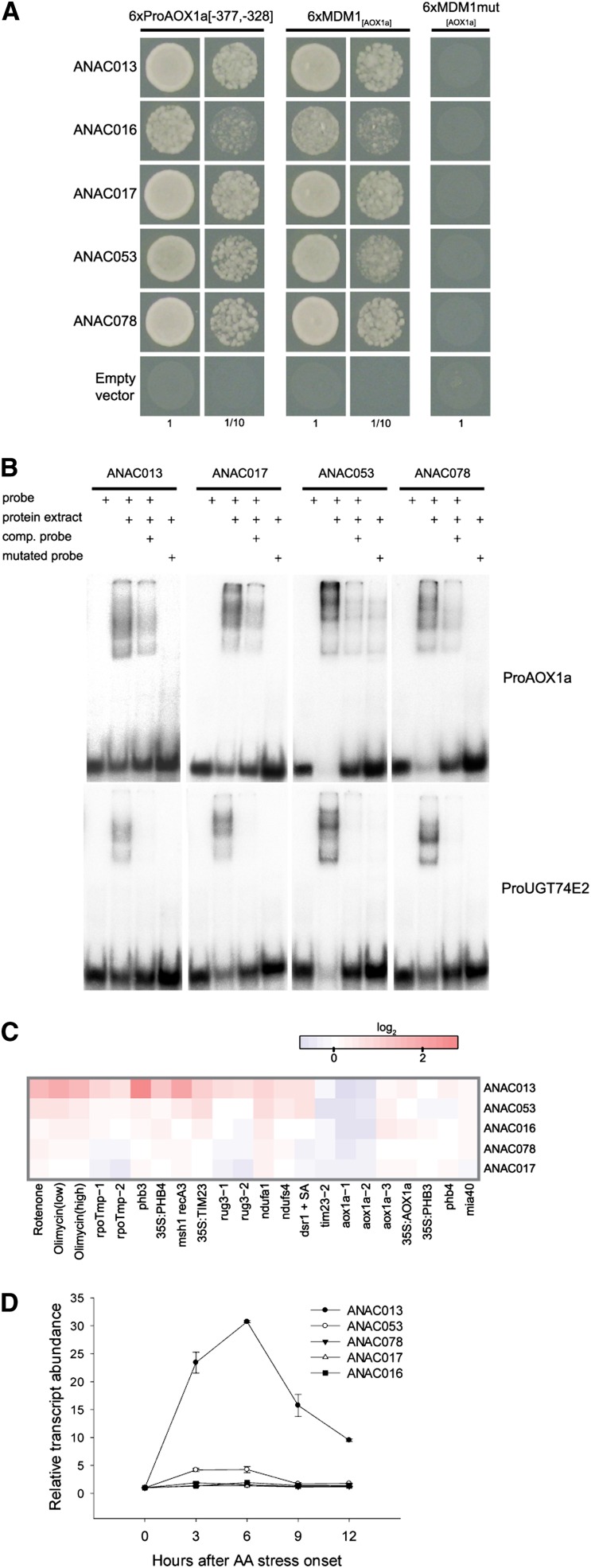
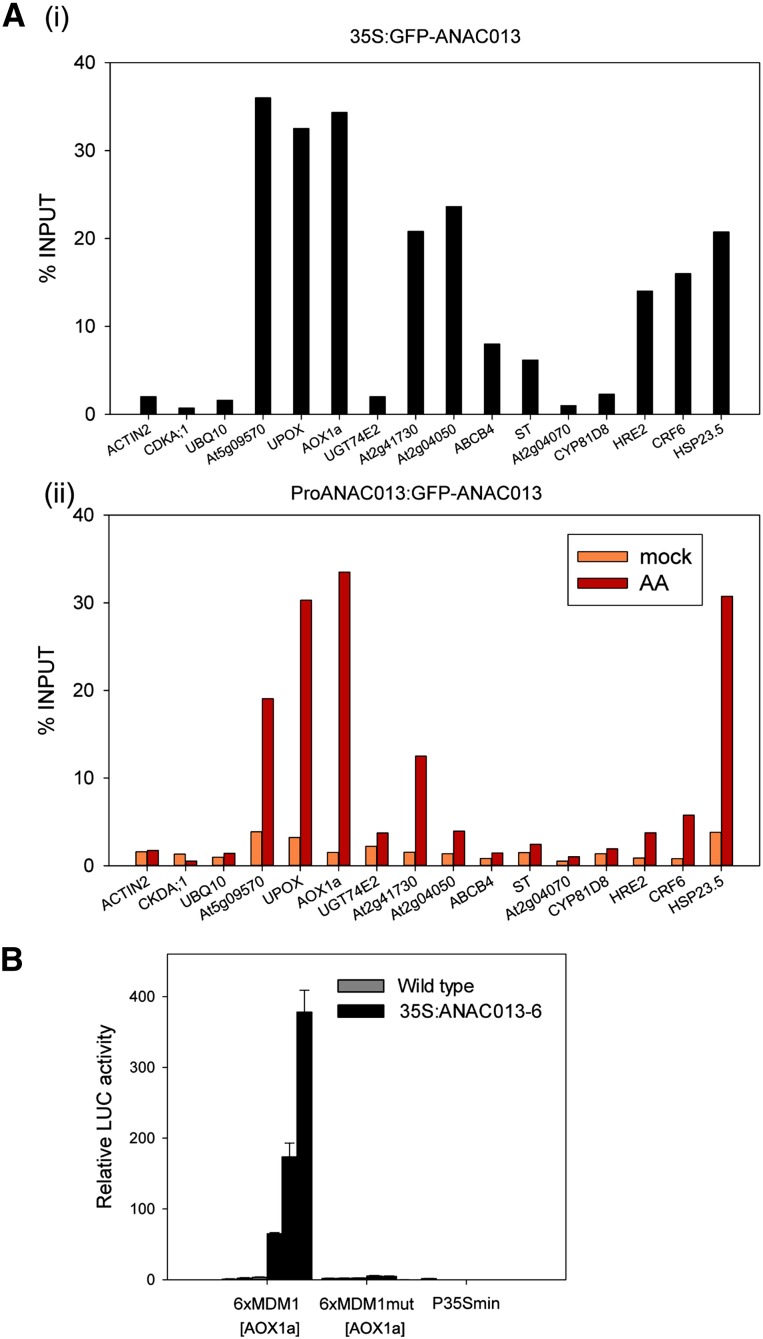
Upon disturbance of their function by stress, mitochondria can signal to the nucleus to steer the expression of responsive genes. This mitochondria-to-nucleus communication is often referred to as mitochondrial retrograde regulation (MRR). Although reactive oxygen species and calcium are likely candidate signaling molecules for MRR, the protein signaling components in plants remain largely unknown. Through meta-analysis of transcriptome data, we detected a set of genes that are common and robust targets of MRR and used them as a bait to identify its transcriptional regulators. In the upstream regions of these mitochondrial dysfunction stimulon (MDS) genes, we found a cis-regulatory element, the mitochondrial dysfunction motif (MDM), which is necessary and sufficient for gene expression under various mitochondrial perturbation conditions. Yeast one-hybrid analysis and electrophoretic mobility shift assays revealed that the transmembrane domain-containing no apical meristem/Arabidopsis transcription activation factor/cup-shaped cotyledon transcription factors (ANAC013, ANAC016, ANAC017, ANAC053, and ANAC078) bound to the MDM cis-regulatory element. We demonstrate that ANAC013 mediates MRR-induced expression of the MDS genes by direct interaction with the MDM cis-regulatory element and triggers increased oxidative stress tolerance. In conclusion, we characterized ANAC013 as a regulator of MRR upon stress in Arabidopsis thaliana.
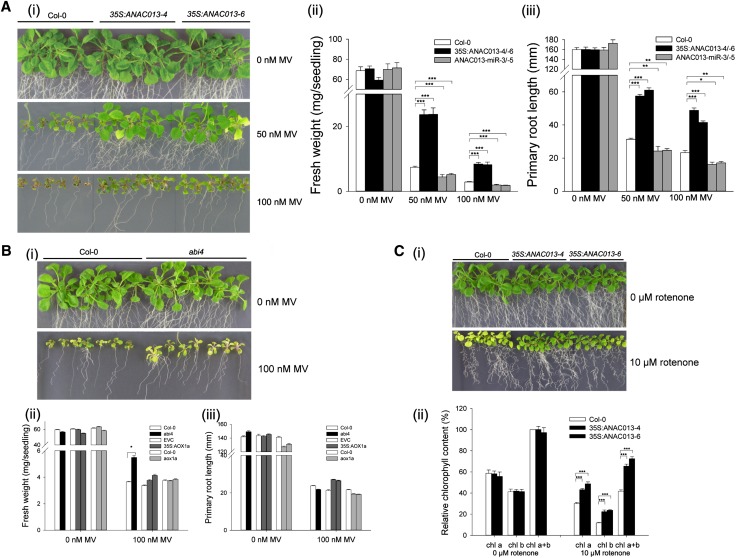
Figures
References
-
- Ahn C.S., Lee J.H., Reum Hwang A., Kim W.T., Pai H.-S. (2006). Prohibitin is involved in mitochondrial biogenesis in plants. Plant J. 46: 658–667 - PubMed
-
- Azevedo D., Tacnet F., Delaunay A., Rodrigues-Pousada C., Toledano M.B. (2003). Two redox centers within Yap1 for H2O2 and thiol-reactive chemicals signaling. Free Radic. Biol. Med. 35: 889–900 - PubMed
-
- Bailey, T.L., and Elkan, C. (1994). Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the Second International Conference on Intelligent Systems for Molecular Biology, R. Altman, D. Brutlag, P. Karp, R. Lathrop, and D. Searls, eds (Menlo Park, CA: AAAI Press), pp. 28–36. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
