

Mouse nasal epithelial innate immune responses to Pseudomonas aeruginosa quorum-sensing molecules require taste signaling components
- PMID: 24045336
- PMCID: PMC4811369
- DOI: 10.1177/1753425913503386
Mouse nasal epithelial innate immune responses to Pseudomonas aeruginosa quorum-sensing molecules require taste signaling components
Abstract
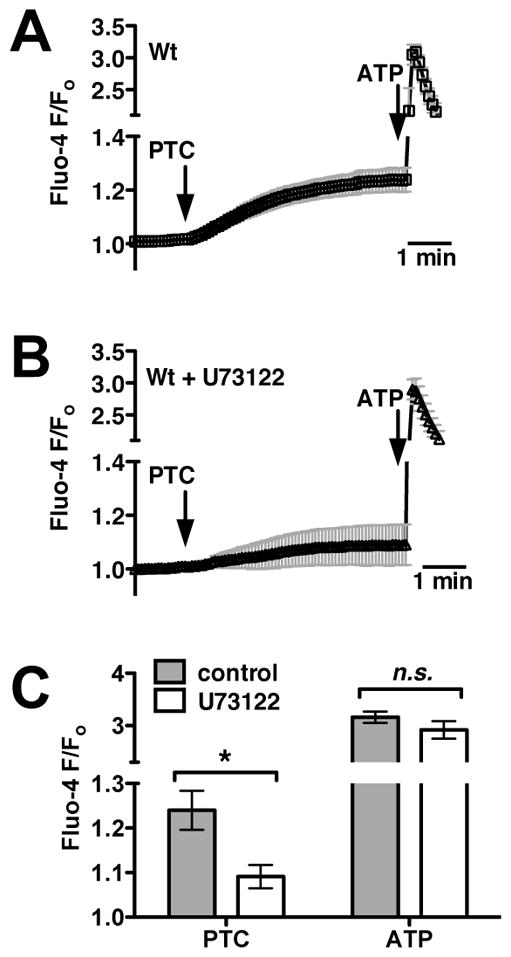
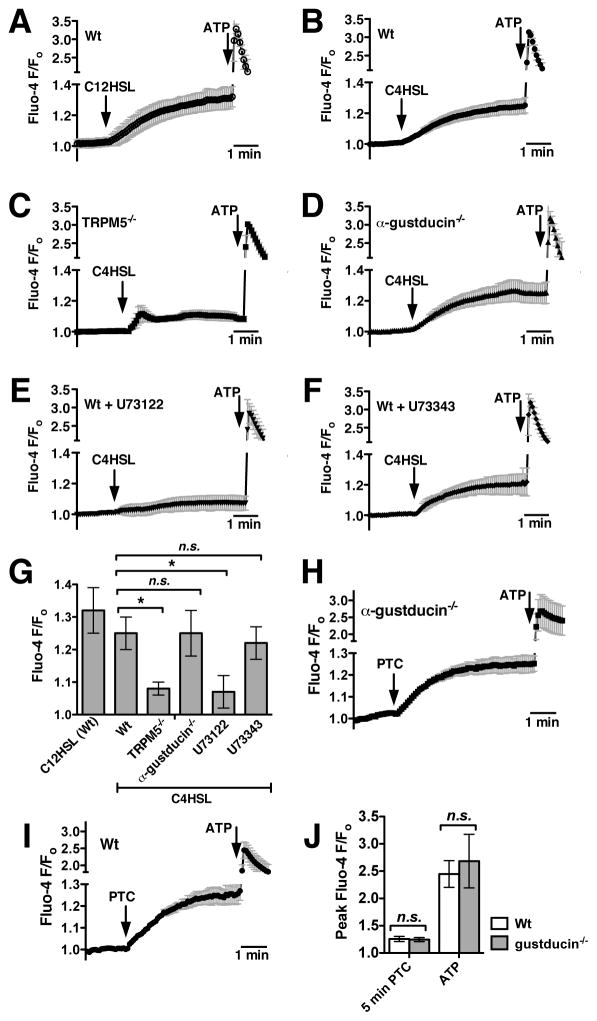
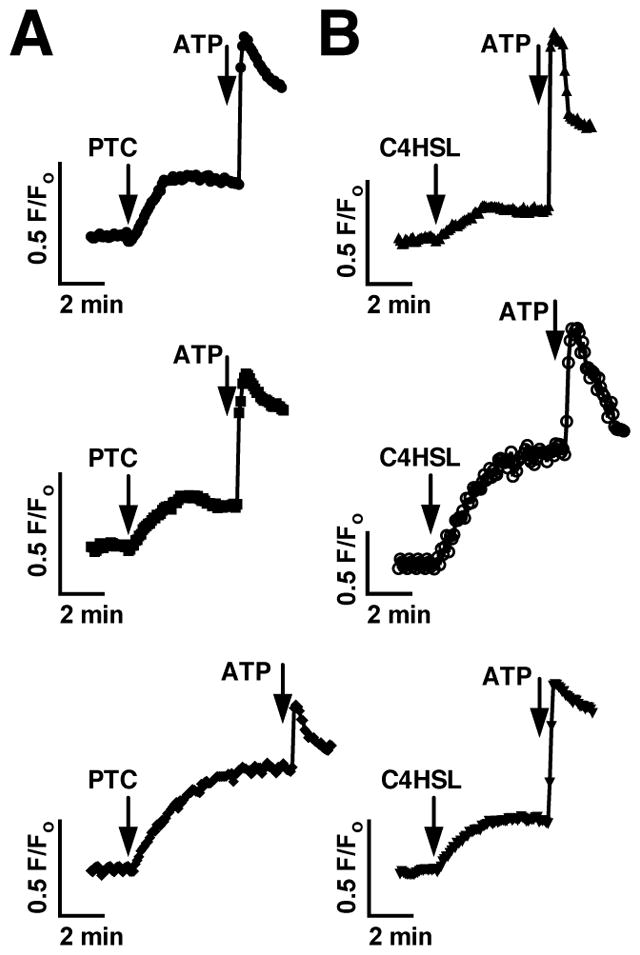
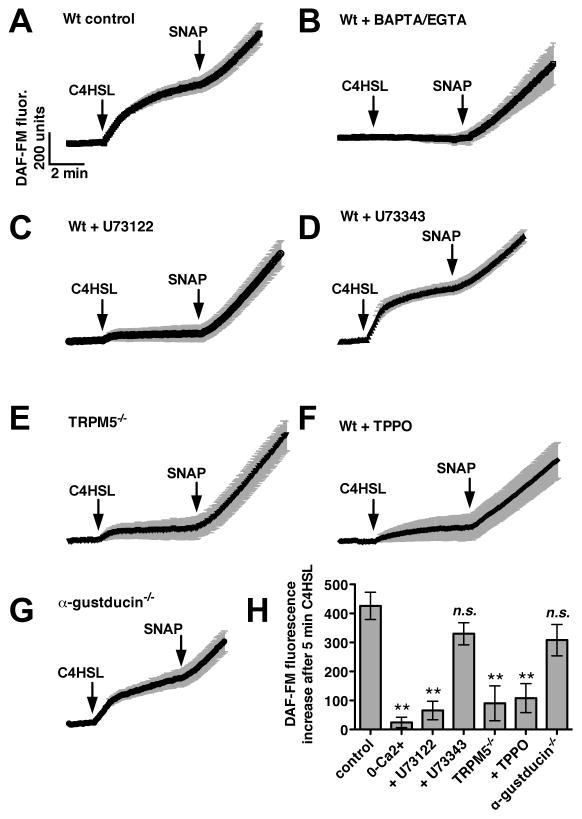
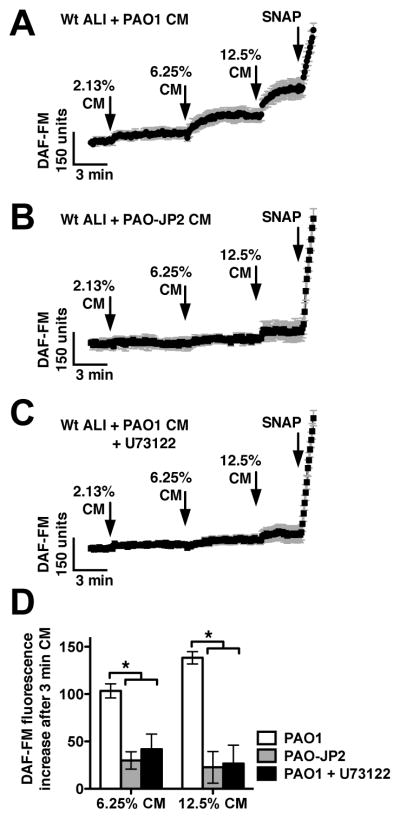
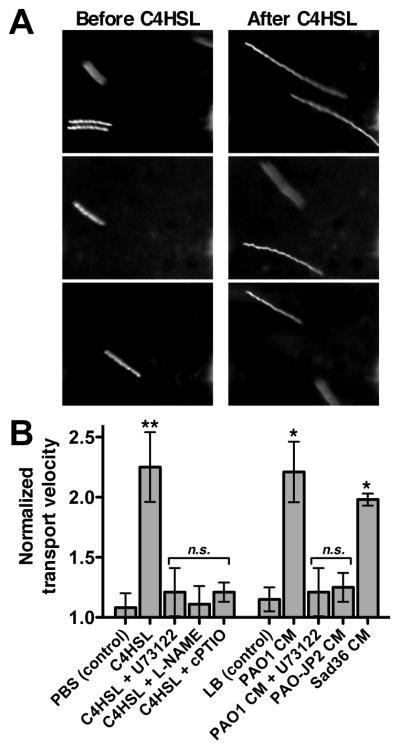
We previously observed that the human bitter taste receptor T2R38 is an important component of upper respiratory innate defense because it detects acyl homoserine lactone (AHL) quorum-sensing molecules secreted by Gram-negative bacteria. T2R38 activation in human sinonasal epithelial cells stimulates calcium and NO signals that increase mucociliary clearance, the major physical respiratory defense against inhaled pathogens. While mice do not have a clear T2R38 ortholog, they do have bitter taste receptors capable of responding to T2R38 agonists, suggesting that T2R-mediated innate immune mechanisms may be conserved in mice. We examined whether AHLs activate calcium and NO signaling in mouse nasal epithelial cells, and utilized pharmacology, as well as cells from knockout mice lacking important components of canonical taste signal transduction pathways, to determine if AHL-stimulated responses require taste signaling molecules. We found that AHLs stimulate calcium-dependent NO production that increases mucociliary clearance and thus likely serves an innate immune role against Gram-negative bacteria. These responses require PLCβ2 and TRPM5 taste signaling components, but not α-gustducin. These data suggest the mouse may be a useful model for further studies of T2R-mediated innate immunity.
Keywords: Acyl-homoserine lactone; T2R bitter taste receptor; chronic rhinosinusitis; innate immunity; mucociliary clearance; nitric oxide.
© The Author(s) 2013 Reprints and permissions: sagepub.co.uk/journalsPermissions.nav.
Conflict of interest statement
The authors have no conflicts of interest to declare pertaining to this article.
Figures
References
-
- Satir P, Sleigh MA. The physiology of cilia and mucociliary interactions. Annu Rev Physiol. 1990;52:137–55. - PubMed
-
- Sleigh MA, Blake JR, Liron N. The propulsion of mucus by cilia. American Rev Respir Dis. 1988;137:726–41. - PubMed
-
- Eliezer N, Sade J, Silberberg A, Nevo AC. The role of mucus in transport by cilia. Am Rev Respir Dis. 1970;102:48–52. - PubMed
-
- Antunes MB, Gudis DA, Cohen NA. Epithelium, cilia, and mucus: their importance in chronic rhinosinusitis. Immunol Allergy Clin North Am. 2009;29:631–43. - PubMed
-
- Cohen NA. Sinonasal mucociliary clearance in health and disease. Ann Otol Rhinol Laryngol Suppl. 2006;196:20–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
