

The adaptor molecule signaling lymphocytic activation molecule (SLAM)-associated protein (SAP) is essential in mechanisms involving the Fyn tyrosine kinase for induction and progression of collagen-induced arthritis
- PMID: 24045941
- PMCID: PMC3814739
- DOI: 10.1074/jbc.M113.473736
The adaptor molecule signaling lymphocytic activation molecule (SLAM)-associated protein (SAP) is essential in mechanisms involving the Fyn tyrosine kinase for induction and progression of collagen-induced arthritis
Abstract
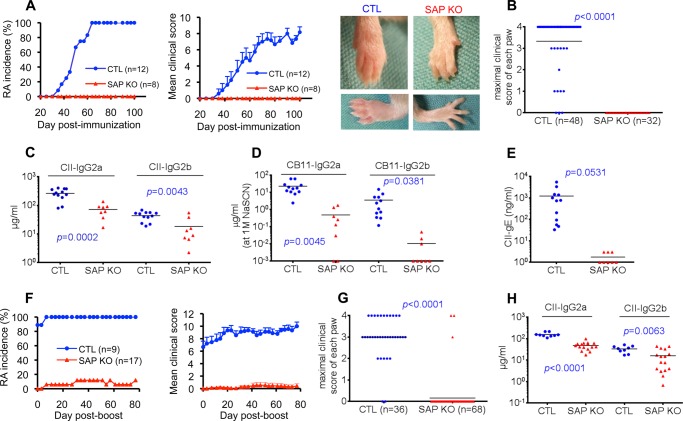
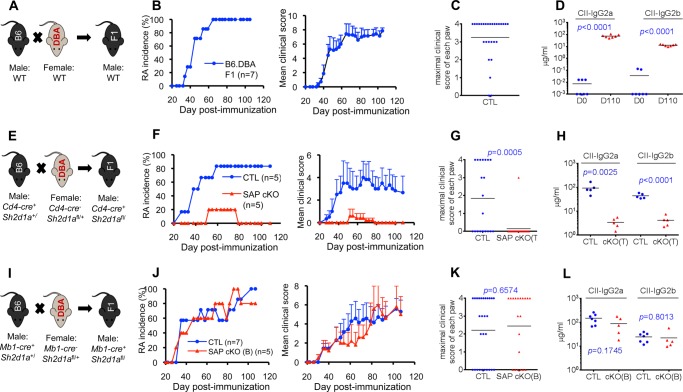
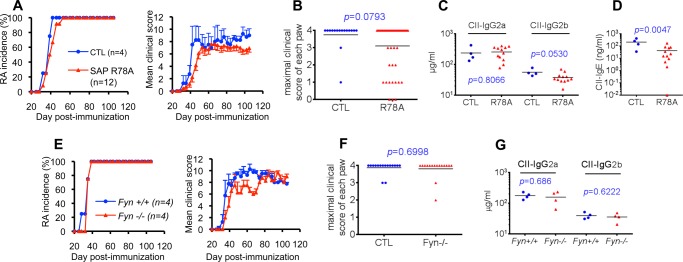
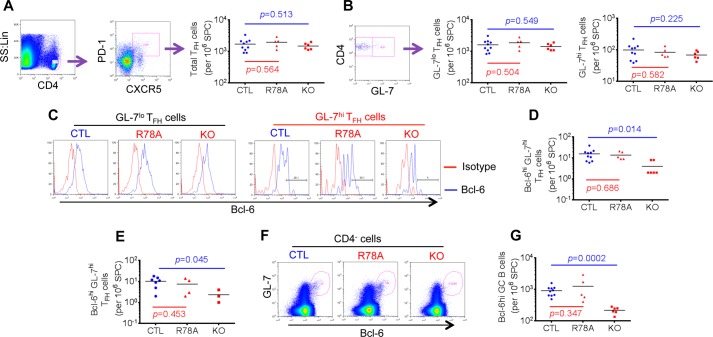
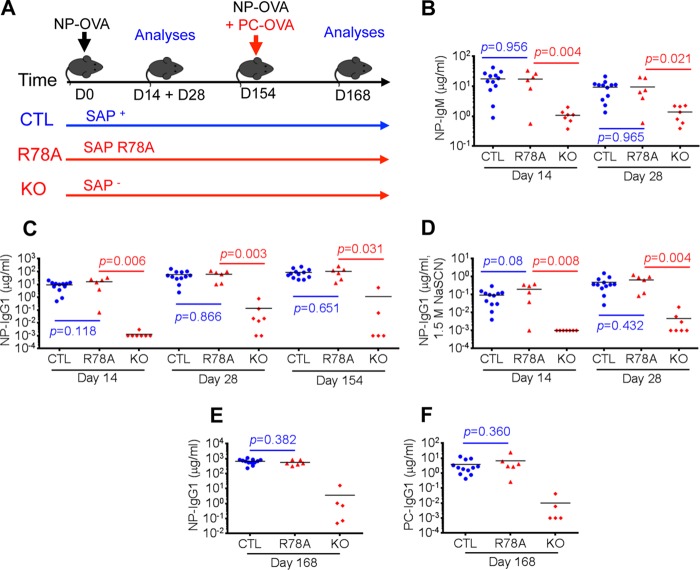
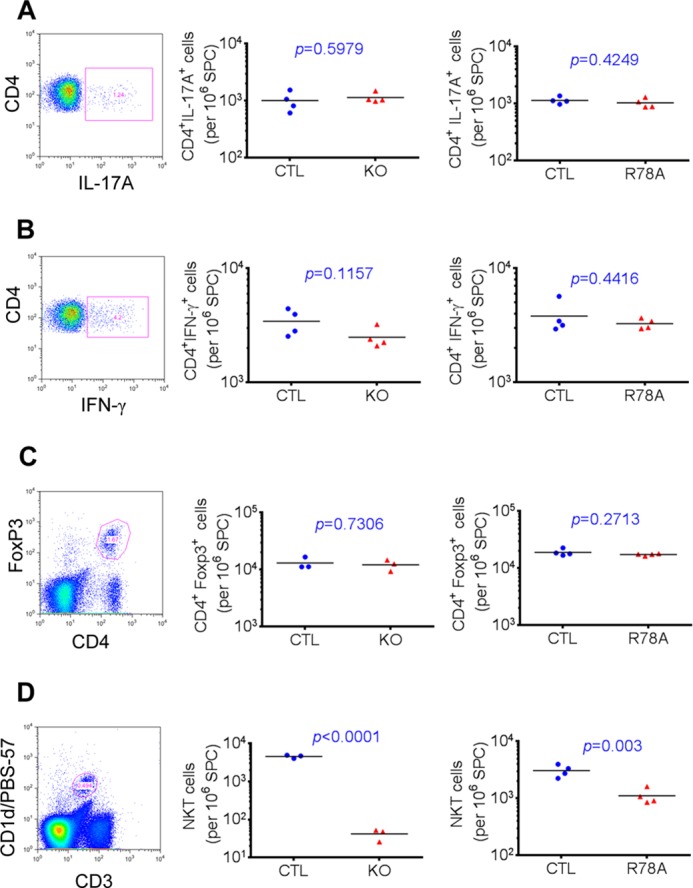
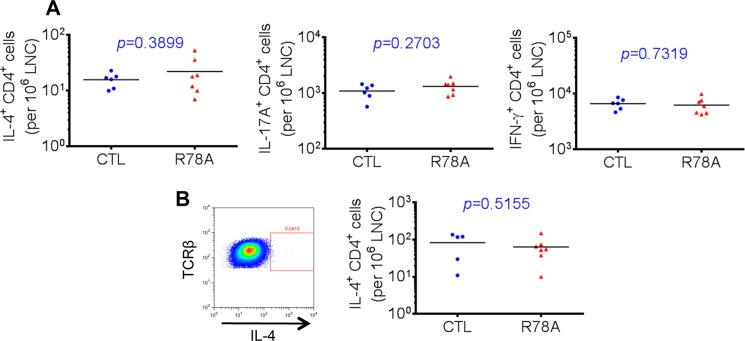
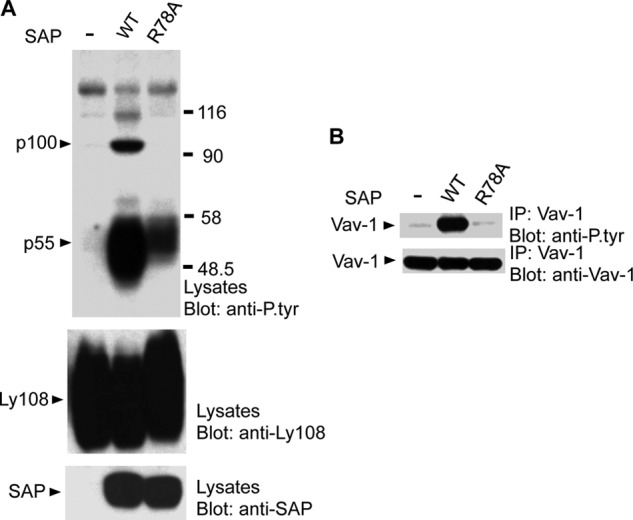
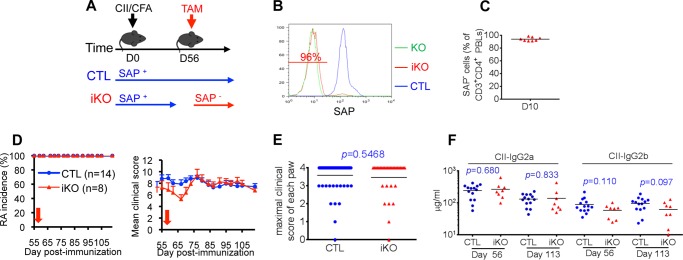
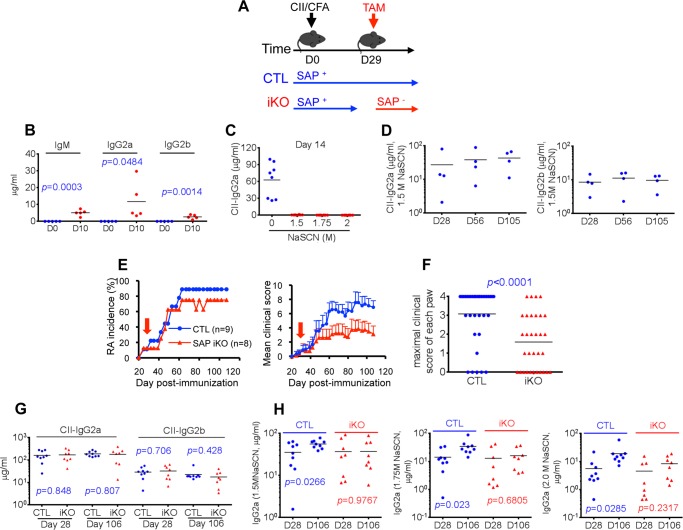
Signaling lymphocytic activation molecule-associated protein (SAP) is an Src homology 2 domain-only adaptor involved in multiple immune cell functions. It has also been linked to immunodeficiencies and autoimmune diseases, such as systemic lupus erythematosus. Here, we examined the role and mechanism of action of SAP in autoimmunity using a mouse model of autoimmune arthritis, collagen-induced arthritis (CIA). We found that SAP was essential for development of CIA in response to collagen immunization. It was also required for production of collagen-specific antibodies, which play a key role in disease pathogenesis. These effects required SAP expression in T cells, not in B cells. In mice immunized with a high dose of collagen, the activity of SAP was nearly independent of its ability to bind the protein tyrosine kinase Fyn and correlated with the capacity of SAP to promote full differentiation of follicular T helper (TFH) cells. However, with a lower dose of collagen, the role of SAP was more dependent on Fyn binding, suggesting that additional mechanisms other than TFH cell differentiation were involved. Further studies suggested that this might be due to a role of the SAP-Fyn interaction in natural killer T cell development through the ability of SAP-Fyn to promote Vav-1 activation. We also found that removal of SAP expression during progression of CIA attenuated disease severity. However, it had no effect on disease when CIA was clinically established. Together, these results indicate that SAP plays an essential role in CIA because of Fyn-independent and Fyn-dependent effects on TFH cells and, possibly, other T cell types.
Keywords: Adaptor Proteins; Autoimmune Diseases; Immunology; SH2 Domains; Src.
Figures

Similar articles
-
Control of T lymphocyte signaling by Ly108, a signaling lymphocytic activation molecule family receptor implicated in autoimmunity.J Biol Chem. 2008 Jul 11;283(28):19255-64. doi: 10.1074/jbc.M800209200. Epub 2008 May 15. J Biol Chem. 2008. PMID: 18482989
-
SAP couples Fyn to SLAM immune receptors.Nat Cell Biol. 2003 Feb;5(2):155-60. doi: 10.1038/ncb920. Nat Cell Biol. 2003. PMID: 12545174
-
Differential requirement for the SAP-Fyn interaction during NK T cell development and function.J Immunol. 2008 Aug 15;181(4):2311-20. doi: 10.4049/jimmunol.181.4.2311. J Immunol. 2008. PMID: 18684920 Free PMC article.
-
Role of SLAM-associated protein in the pathogenesis of autoimmune diseases and immunological disorders.Arch Immunol Ther Exp (Warsz). 2010 Feb;58(1):37-44. doi: 10.1007/s00005-009-0060-7. Epub 2010 Jan 5. Arch Immunol Ther Exp (Warsz). 2010. PMID: 20049647 Review.
-
SLAM-SAP-Fyn: Old Players with New Roles in iNKT Cell Development and Function.Int J Mol Sci. 2019 Sep 27;20(19):4797. doi: 10.3390/ijms20194797. Int J Mol Sci. 2019. PMID: 31569599 Free PMC article. Review.
Cited by
-
Identification of predictors for bacterial meningitis diagnosis based on transcriptomics and genetic analysis.AMB Express. 2025 May 30;15(1):84. doi: 10.1186/s13568-025-01893-7. AMB Express. 2025. PMID: 40445403 Free PMC article.
-
Decreased SAP Expression in T Cells from Patients with Systemic Lupus Erythematosus Contributes to Early Signaling Abnormalities and Reduced IL-2 Production.J Immunol. 2016 Jun 15;196(12):4915-24. doi: 10.4049/jimmunol.1501523. Epub 2016 May 11. J Immunol. 2016. PMID: 27183584 Free PMC article.
-
Signal Transducer and Activator of Transcription 3 Hyperactivation Associates With Follicular Helper T Cell Differentiation and Disease Activity in Rheumatoid Arthritis.Front Immunol. 2018 Jun 4;9:1226. doi: 10.3389/fimmu.2018.01226. eCollection 2018. Front Immunol. 2018. PMID: 29915585 Free PMC article.
-
A hematopoietic cell-driven mechanism involving SLAMF6 receptor, SAP adaptors and SHP-1 phosphatase regulates NK cell education.Nat Immunol. 2016 Apr;17(4):387-96. doi: 10.1038/ni.3369. Epub 2016 Feb 15. Nat Immunol. 2016. PMID: 26878112
-
Progression of AITL-like tumors in mice is driven by Tfh signature proteins and T-B cross talk.Blood Adv. 2020 Mar 10;4(5):868-879. doi: 10.1182/bloodadvances.2019001114. Blood Adv. 2020. PMID: 32130407 Free PMC article.
References
-
- Schwartzberg P. L., Mueller K. L., Qi H., Cannons J. L. (2009) SLAM receptors and SAP influence lymphocyte interactions, development and function. Nat. Rev. Immunol. 9, 39–46 - PubMed
-
- Veillette A., Dong Z., Latour S. (2007) Consequence of the SLAM-SAP signaling pathway in innate-like and conventional lymphocytes. Immunity 27, 698–710 - PubMed
-
- Calpe S., Wang N., Romero X., Berger S. B., Lanyi A., Engel P., Terhorst C. (2008) The SLAM and SAP gene families control innate and adaptive immune responses. Adv. Immunol. 97, 177–250 - PubMed
-
- Ma C. S., Nichols K. E., Tangye S. G. (2007) Regulation of cellular and humoral immune responses by the SLAM and SAP families of molecules. Annu. Rev. Immunol. 25, 337–379 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
