

Val66Met polymorphism of BDNF alters prodomain structure to induce neuronal growth cone retraction
- PMID: 24048383
- PMCID: PMC3820160
- DOI: 10.1038/ncomms3490
Val66Met polymorphism of BDNF alters prodomain structure to induce neuronal growth cone retraction
Erratum in
- Nat Commun. 2014;5:3564
Abstract
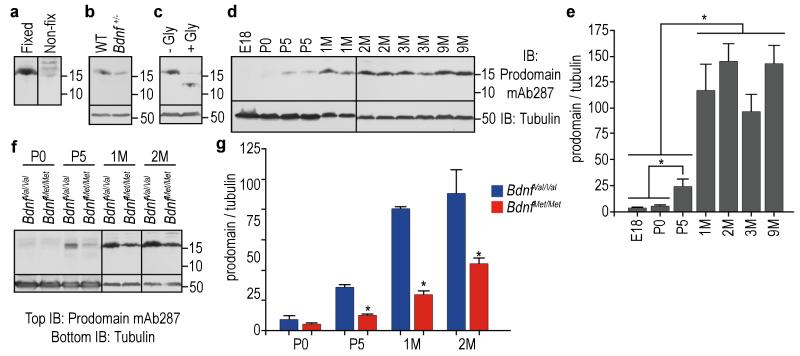
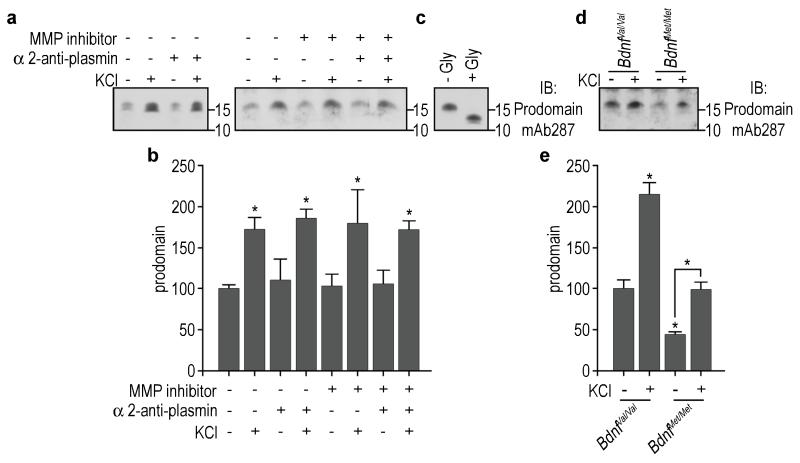
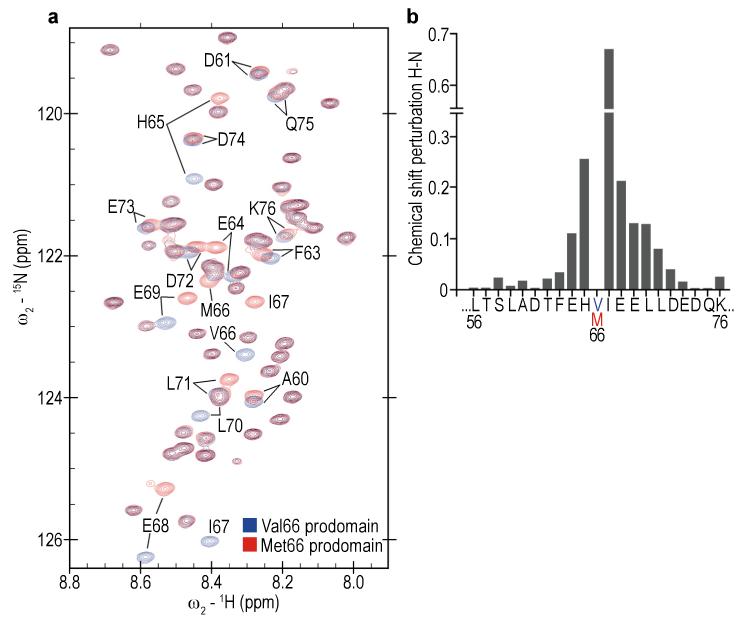
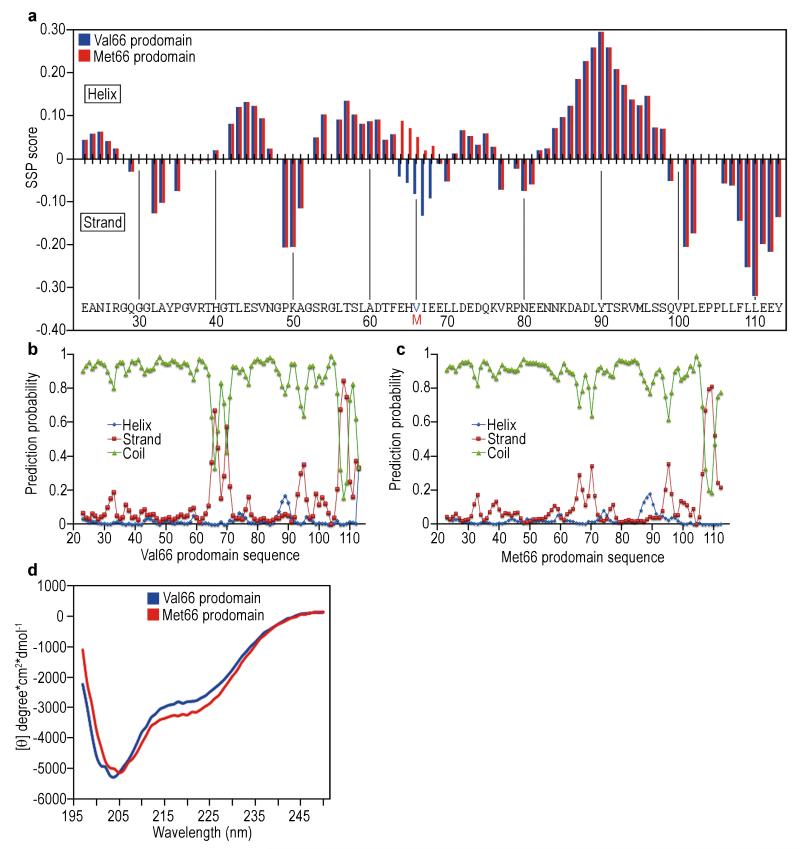
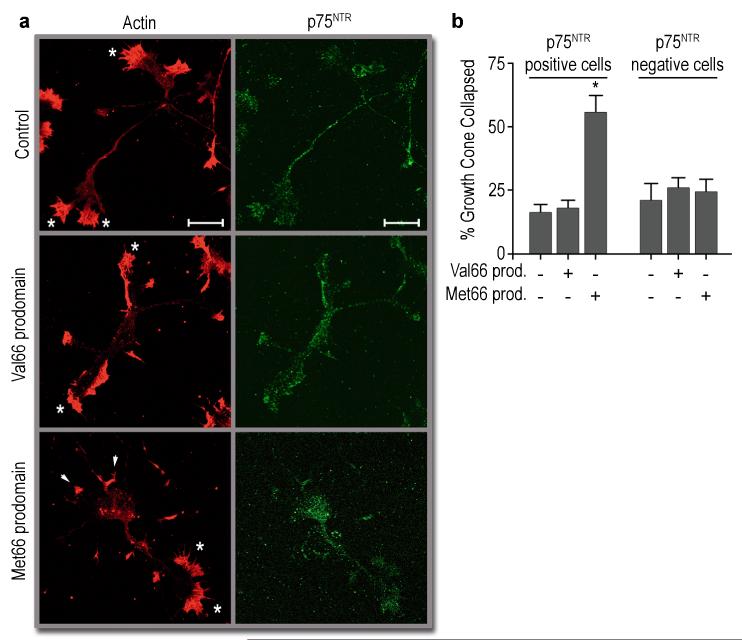
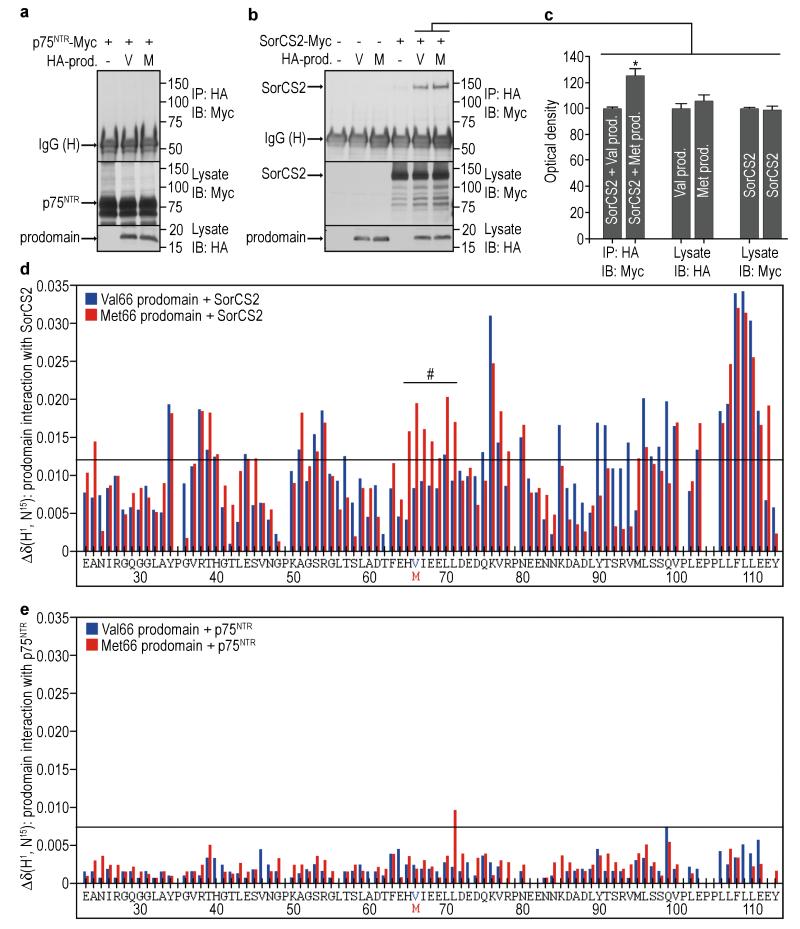
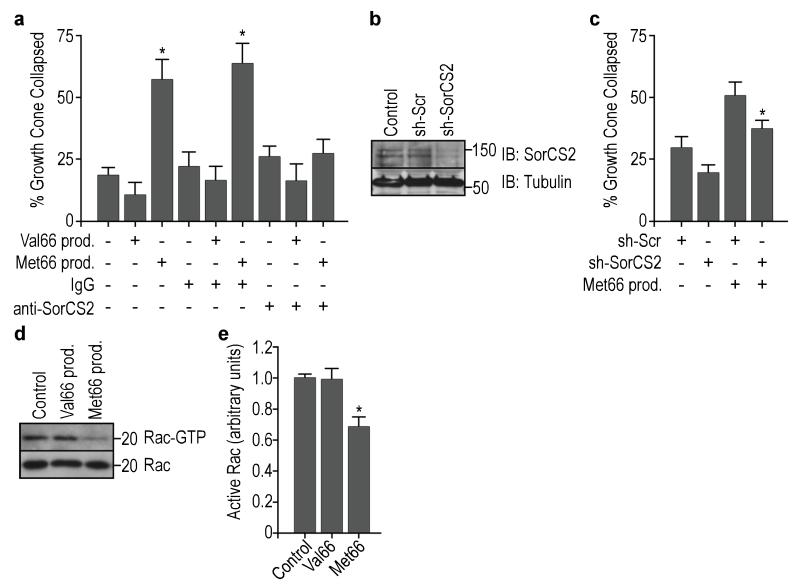
A common single-nucleotide polymorphism (SNP) in the human brain-derived neurotrophic factor (BDNF) gene results in a Val66Met substitution in the BDNF prodomain region. This SNP is associated with alterations in memory and with enhanced risk to develop depression and anxiety disorders in humans. Here we show that the isolated BDNF prodomain is detected in the hippocampus and that it can be secreted from neurons in an activity-dependent manner. Using nuclear magnetic resonance spectroscopy and circular dichroism, we find that the prodomain is intrinsically disordered, and the Val66Met substitution induces structural changes. Surprisingly, application of Met66 (but not Val66) BDNF prodomain induces acute growth cone retraction and a decrease in Rac activity in hippocampal neurons. Expression of p75(NTR) and differential engagement of the Met66 prodomain to the SorCS2 receptor are required for this effect. These results identify the Met66 prodomain as a new active ligand, which modulates neuronal morphology.
Figures
References
-
- Egan MF, et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell. 2003;112:257–269. - PubMed
-
- Verhagen M, et al. Meta-analysis of the BDNF Val66Met polymorphism in major depressive disorder: effects of gender and ethnicity. Mol Psychiatry. 2010;15:260–271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10-RR023694-01EWOF/RR/NCRR NIH HHS/United States
- P01 HD023315/HD/NICHD NIH HHS/United States
- R01 NS064114/NS/NINDS NIH HHS/United States
- NS21072/NS/NINDS NIH HHS/United States
- R01 NS021072/NS/NINDS NIH HHS/United States
- S10 RR023694/RR/NCRR NIH HHS/United States
- R01 NS052819/NS/NINDS NIH HHS/United States
- HD23315/HD/NICHD NIH HHS/United States
- R01 NS030687/NS/NINDS NIH HHS/United States
- R01 AG025970/AG/NIA NIH HHS/United States
- NS030687/NS/NINDS NIH HHS/United States
- R56 NS021072/NS/NINDS NIH HHS/United States
- NS064114/NS/NINDS NIH HHS/United States
- NS052819/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
