

Fibroblast growth factor 21 is not required for the antidiabetic actions of the thiazoladinediones
- PMID: 24049735
- PMCID: PMC3773835
- DOI: 10.1016/j.molmet.2013.05.005
Fibroblast growth factor 21 is not required for the antidiabetic actions of the thiazoladinediones
Abstract
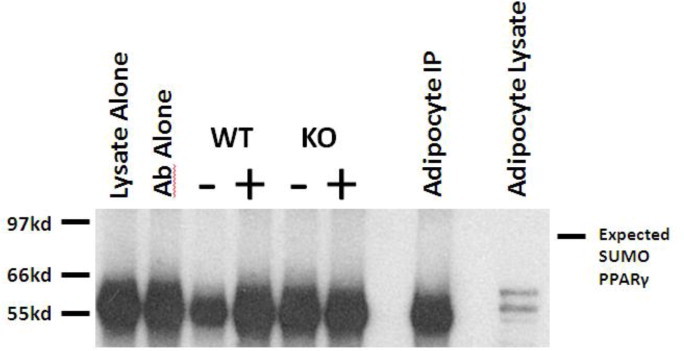
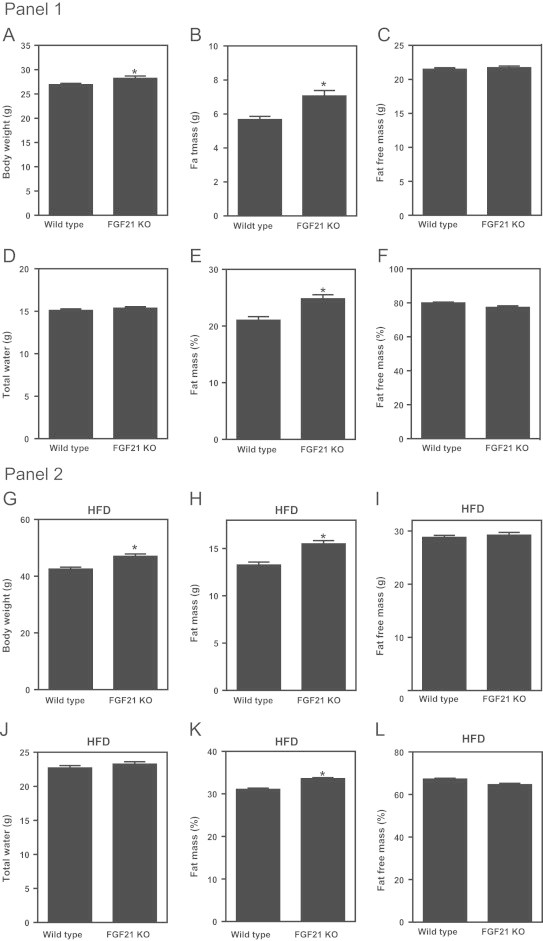
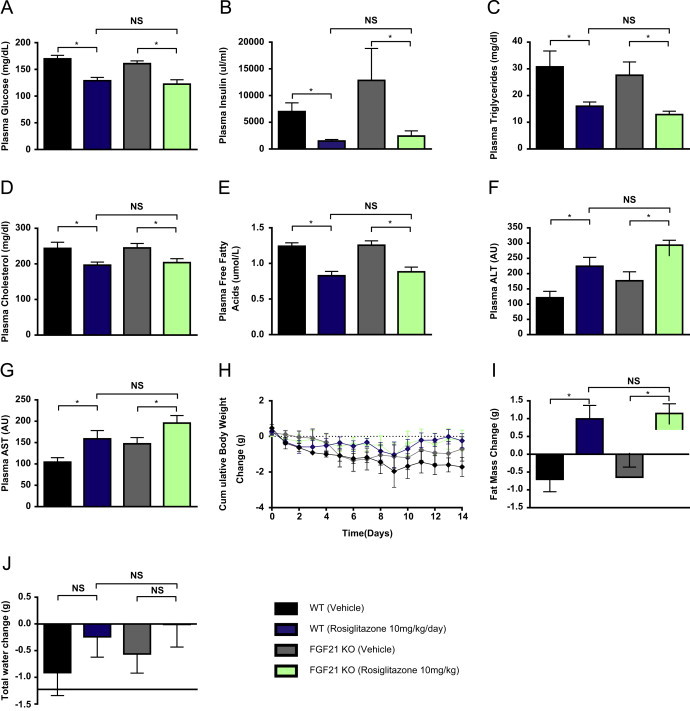
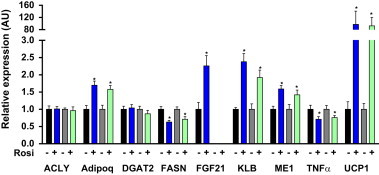
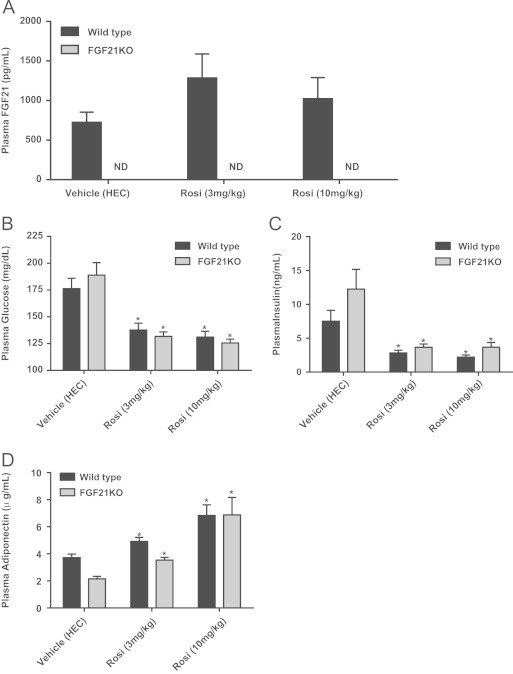
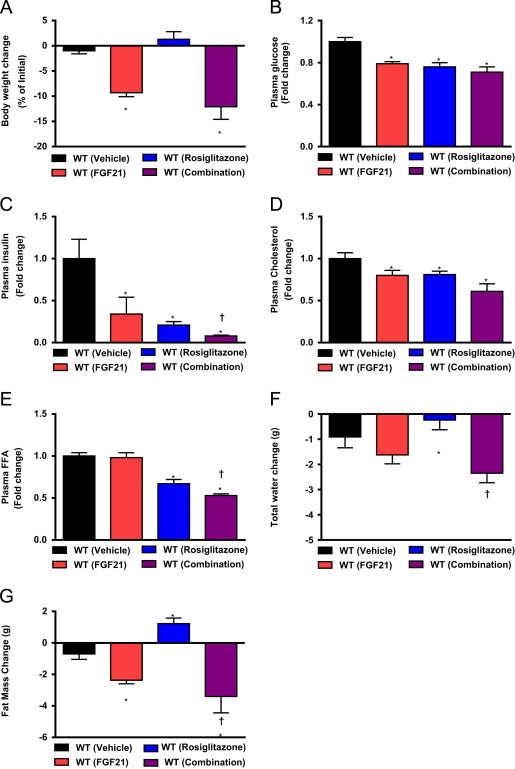
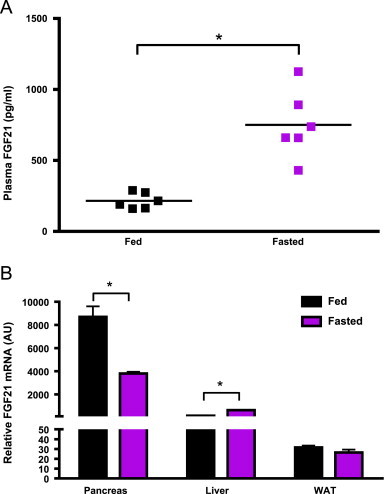
Fibroblast growth factor 21 is an emerging metabolic regulator that was recently proposed to be a fed-state inducible factor in adipose tissue. As mice lacking FGF21 were refractory to treatment with rosiglitazone, FGF21 was suggested to underlie PPARγ-driven pharmacology and side effect profile (Dutchak et al., 2012 [12]). To evaluate FGF21/PPARγ cross-talk we conducted experiments in control and FGF21 null animals and found that rosiglitazone was equally efficacious in both strains. Specifically, diverse endpoints ranging from enhanced glycemic control, improved lipid homeostasis and side effects such as adipose accumulation were evident in both genotypes. Furthermore, the transcriptional signature and cytokine secretion profile of rosiglitazone action were maintained in our FGF21KO animals. Finally, we found that FGF21 in adipose was expressed at comparable levels in fasted and fed states. Thus, our data present a new viewpoint on the FGF21/PPARγ interplay whereby FGF21 is not necessary for the metabolic events downstream of PPARγ.
Keywords: Adiponectin; FGF21; Metabolism; PPARγ; Rosiglitazone.
Figures
References
-
- Adams A.C., Kharitonenkov A. FGF21: the center of a transcriptional nexus in metabolic regulation. Current Diabetes Reviews. 2012;8:285–293. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
