

Control of transcriptional elongation
- PMID: 24050178
- PMCID: PMC3974797
- DOI: 10.1146/annurev-genet-110711-155440
Control of transcriptional elongation
Abstract
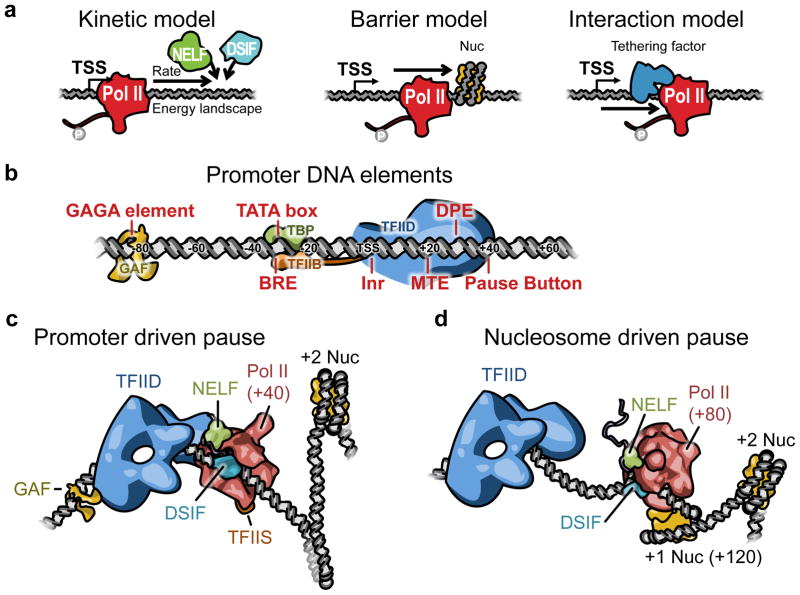
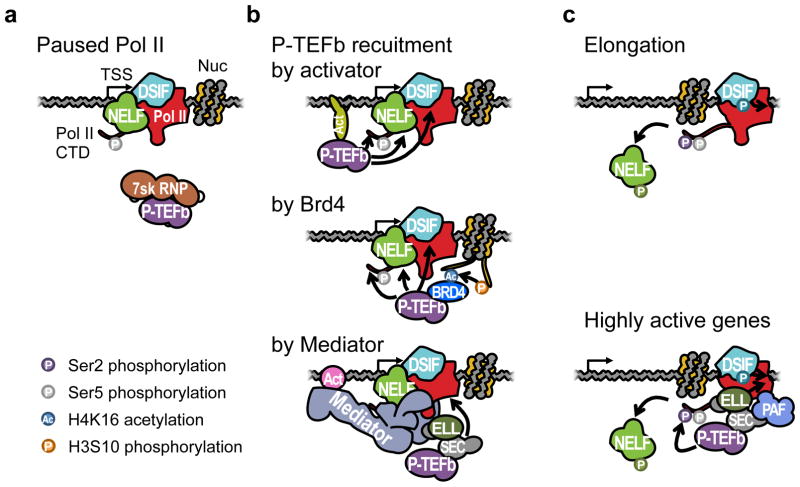
Elongation is becoming increasingly recognized as a critical step in eukaryotic transcriptional regulation. Although traditional genetic and biochemical studies have identified major players of transcriptional elongation, our understanding of the importance and roles of these factors is evolving rapidly through the recent advances in genome-wide and single-molecule technologies. Here, we focus on how elongation can modulate the transcriptional outcome through the rate-liming step of RNA polymerase II (Pol II) pausing near promoters and how the participating factors were identified. Among the factors we describe are the pausing factors--NELF (negative elongation factor) and DSIF (DRB sensitivity-inducing factor)--and P-TEFb (positive elongation factor b), which is the key player in pause release. We also describe the high-resolution view of Pol II pausing and propose nonexclusive models for how pausing is achieved. We then discuss Pol II elongation through the bodies of genes and the roles of FACT and SPT6, factors that allow Pol II to move through nucleosomes.
Figures


References
Literature Cited
-
- Adelman K, Marr MT, Werner J, Saunders A, Ni Z, Andrulis ED, Lis JT. Efficient release from promoter-proximal stall sites requires transcript cleavage factor TFIIS. Mol Cell. 2005;17(1):103–112. - PubMed
-
- Ardehali MB, Lis JT. Tracking rates of transcription and splicing in vivo. Nat Struct Mol Biol. 2009;16(11):1123–1124. - PubMed
-
- Aso T, Lane W, Conaway J, Conaway R. Elongin (SIII): a multisubunit regulator of elongation by RNA polymerase II. Science. 1995;269(5229):1439–1443. - PubMed
Related Resources
-
- Bermejo R, Lai MS, Foiani M. Preventing replication stress to maintain genome stability: resolving conflicts between replication and transcription. Mol Cell. 2012;45(6):710–718. - PubMed
-
- Selth LA, Sigurdsson S, Svejstrup JQ. Transcript Elongation by RNA Polymerase II. Annu Rev Biochem. 2010;79:271–293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
