

Review
doi: 10.1007/s00726-013-1593-y.
Epub 2013 Sep 20.
GPCR: G protein complexes--the fundamental signaling assembly
Affiliations
- PMID: 24052187
- PMCID: PMC3845202
- DOI: 10.1007/s00726-013-1593-y
Item in Clipboard
Review
GPCR: G protein complexes--the fundamental signaling assembly
Amino Acids.
2013 Dec.
Abstract
G protein coupled receptors (GPCR) constitute the largest group of cell surface receptors that transmit various signals across biological membranes through the binding and activation of heterotrimeric G proteins, which amplify the signal and activate downstream effectors leading to the biological responses. Thus, the first critical step in this signaling cascade is the interaction between receptor and its cognate G protein. Understanding this critical event at the molecular level is of high importance because abnormal function of GPCRs is associated with many diseases. Thus, these receptors are targets for drug development.
Conflict of interest statement
The author declares that she has no conflict of interst.
Figures
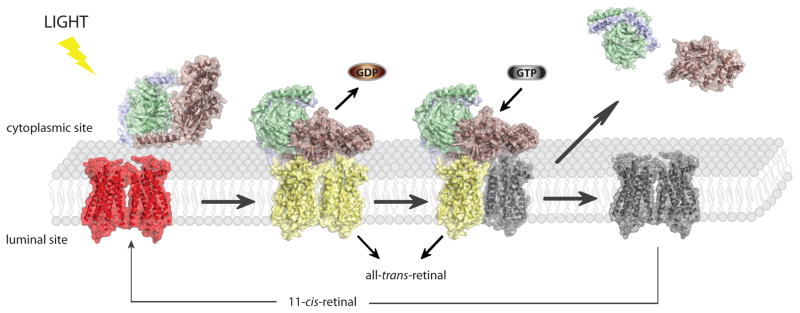
Rhodopsin signaling. Light activation of rhodopsin enables binding of its cognate G protein, transducin and triggers GDP→GTP exchange in its α subunit. Transiently, high affinity, nucleotide free rhodopsin-Gt complex is formed. Binding of GTP causes complex dissociation as well as dissociation of Gtα subunit from Gtβγ dimer and they activate different downstream pathways leading in effect to neuronal response in the brain. Dissociation of transducin from activated rhodopsin enables dissociation of is chromophore all-trans-retinal. Accumulated free opsin has to be regenerated with 11-cis-retinal in order to restore its function.
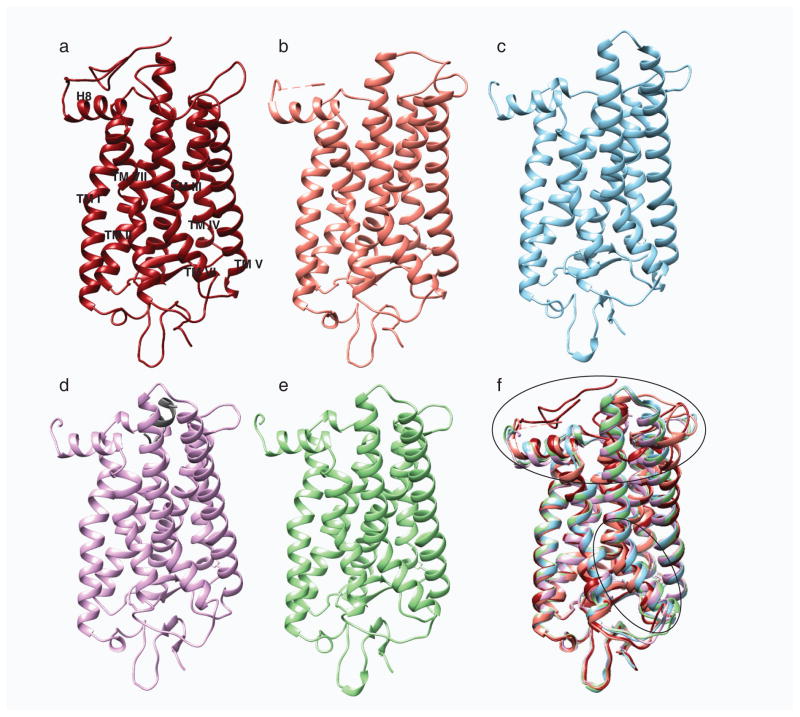
Structural comparison of different photo-states of rhodopsin. a, Structures of ground state rhodopsin (1u19) (Pescitelli et al. 2008); b, photoactivated rhodopsin (2i35) (Salom et al. 2006); c, opsin (3cap) (Park et al. 2008); d, opsin with bound C-terminal peptide of Gt (3dqb) (Scheerer et al. 2008) and e, Meta II (3pxo) (Choe et al. 2011) are shown in dark red, salmon, blue, purple and green, respectively. f, All activated rhodopsin states were superposed on its ground state structure. Superposition of these structures show rearrangement of the cytoplasmic loops and movement of helix TM VI depicted with black ovals.
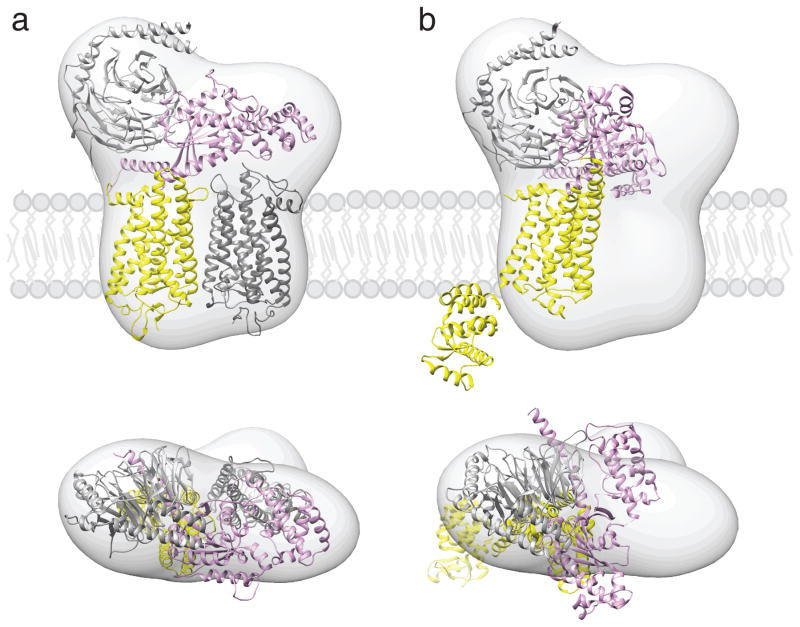
Proposed location of GPCR-G protein complexes in a phospholipid bilayer. Semi-empirical model of the complex formed between light-activated rhodopsin dimer and a Gt heterotrimer (a) (Jastrzebska et al. 2011) and a structure of T4L-β2AR-Gs complex (b) (Rasmussen et al. 2011b) were fitted into a 3D molecular map derived from EM-single particle reconstructions of negatively stained, crosslinked rhodopsin-Gt complexes purified in LMNG. Best fitting of the rhodopsin-Gt model was achieved for rhodopsin dimer and Gt hereotrimer (GOT1) after 30° hinge-like motion of the α-helical domain of Gt was applied. Fitting the T4L-β2AR-Gs-nanobody structure into our EM 3D map leaves enough unoccupied space to accommodate a second molecule of this receptor, conformation of Gsα helical domain is inconsistent with our EM-density. The photoactivated rhodopsin that binds C-terminus of Gtα and molecule of T4L-β2AR is depicted with yellow. The second rhodopsin molecule is shown in dark grey. Gα is colored pink; Gβ and Gγ are colored light grey.
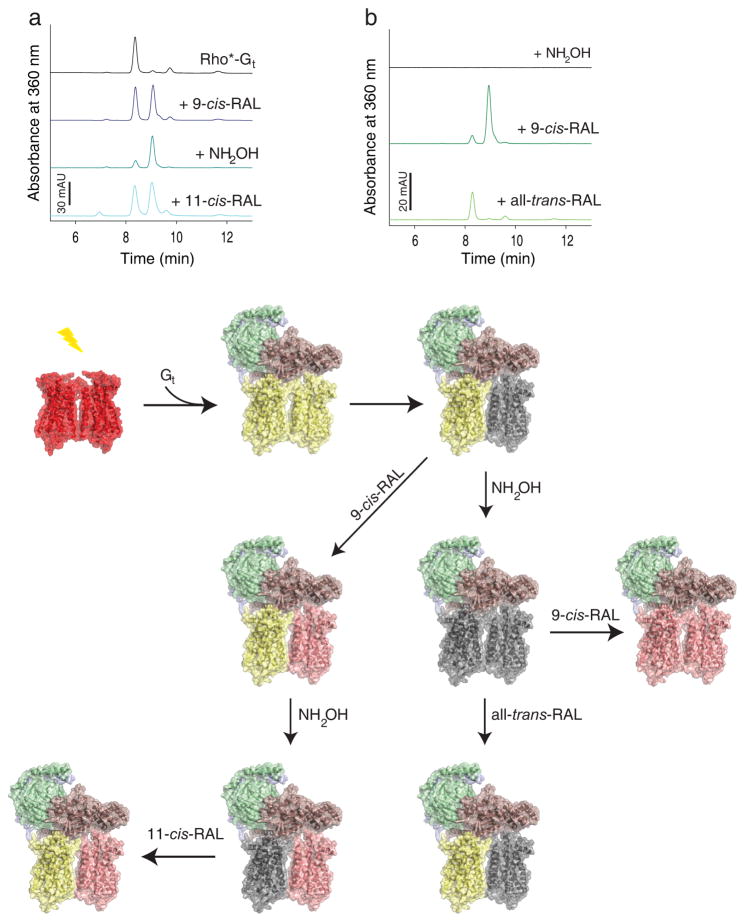
Asymmetry of activated rhodopsin (Rho*) dimer within rhodopsin-Gt (Rho*-Gt) complex. Gt binds to fully light activated rhodopsin (Rho) forming the Rho*-Gt complex. After light illumination, 11-cis-retinal (11-cis-RAL) of ground state rhodopsin (shown as red-red dimer) isomerizes to all-trans-retinal (all-trans-RAL) (yellow-yellow dimer) allowing binding of the Gt heterotrimer (Gtα is depicted in salmon, Gtβ in green and Gtγ in light blue). This binding prevents chromophore release from one rhodopsin in the dimer, introducing dimer asymmetry in the Rho*-Gt complex (shown as a yellow-grey dimer bound to Gt and black trace on the chromatogram). a, retinal-free rhodopsin molecule of the Rho*-Gt complex can be regenerated with 9-cis-retinal (9-cis-RAL) (shown as yellow-pink rhodopsin dimer bound to Gt and blue trays on the chromatogram). Release of chromophore from the Gt-protected rhodopsin molecule can be promoted by NH2OH, resulting then in formation of an asymmetric retinal free and 9-cis-retinal regenerated rhodopsin dimer complexed to Gt (shown as grey-pink rhodopsin dimer bound to Gt and dark cyan trace on the chromatogram). Regeneration of this complex with 11-cis-retinal surprisingly results in formation of all-trans-retinal bound and 9-cis-retinal bound asymmetric rhodopsin dimer in the complex with Gt (shown as yellow-pink rhodopsin dimer bound to Gt and cyan trace on the chromatogram). b, the chromophore-free Rho*-Gt complex (modeled as a grey-grey rhodopsin dimer bound to Gt and black lane on the chromatogram) can be generated by treatment of the Rho*-Gt complex with NH2OH. Then this chromophore-free Rho*-Gt complex can be regenerated with 9-cis-retinal which binds to both rhodopsin monomers within the dimer (shown as a pink-pink dimer bound to Gt and dark green trace on the chromatogram). Alternatively, it can be regenerated with all-trans-retinal but only one rhodopsin molecule, the one directly coupled to Gt binds all-trans-retinal (shown as a yellow-grey dimer bound to Gt and light green trace on the chromatogram).
References
-
- Albizu L, Cottet M, Kralikova M, Stoev S, Seyer R, Brabet I, Roux T, Bazin H, Bourrier E, Lamarque L, Breton C, Rives ML, Newman A, Javitch J, Trinquet E, Manning M, Pin JP, Mouillac B, Durroux T. Time-resolved FRET between GPCR ligands reveals oligomers in native tissues. Nat Chem Biol. 2010;6 (8):587–594. doi: 10.1038/nchembio.396. - DOI - PMC - PubMed
-
- Baneres JL, Parello J. Structure-based analysis of GPCR function: evidence for a novel pentameric assembly between the dimeric leukotriene B4 receptor BLT1 and the G-protein. Journal of molecular biology. 2003;329 (4):815–829. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
