

STING-IRF3 pathway links endoplasmic reticulum stress with hepatocyte apoptosis in early alcoholic liver disease
- PMID: 24052526
- PMCID: PMC3799324
- DOI: 10.1073/pnas.1308331110
STING-IRF3 pathway links endoplasmic reticulum stress with hepatocyte apoptosis in early alcoholic liver disease
Abstract
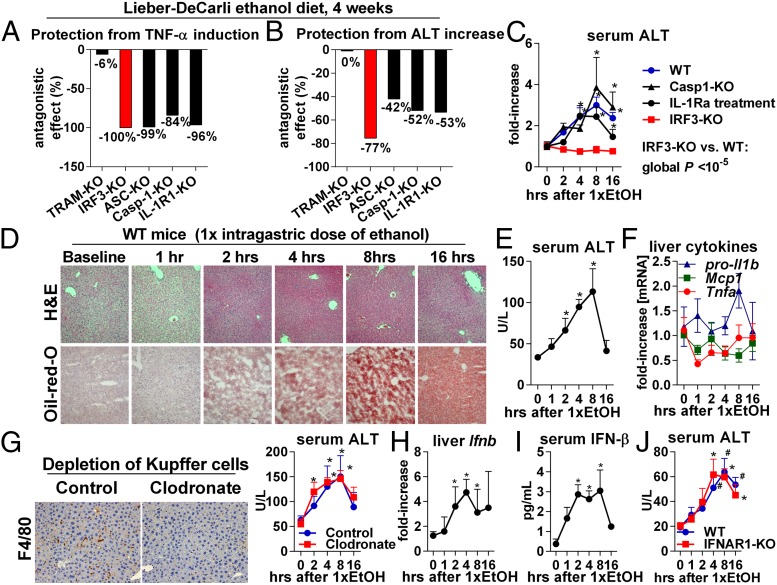
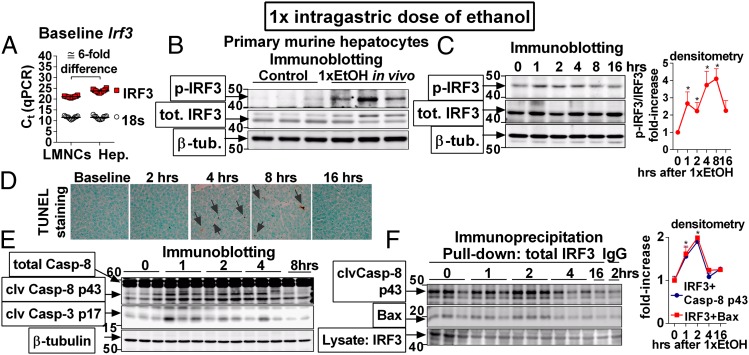
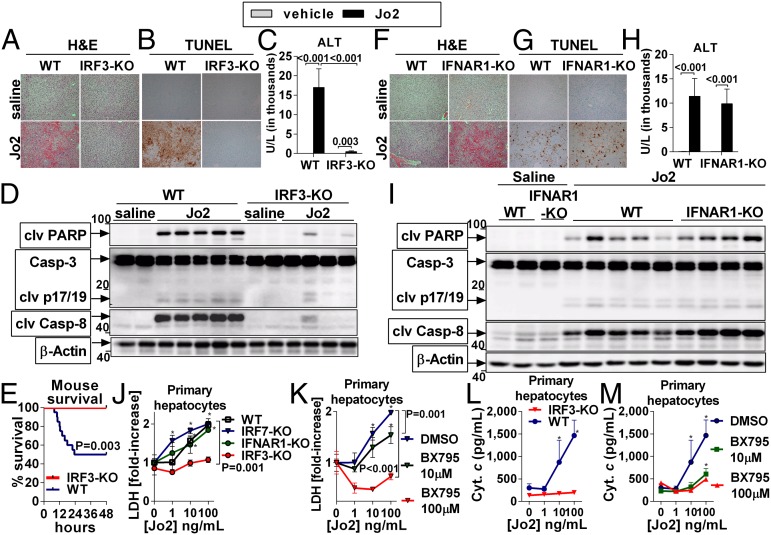
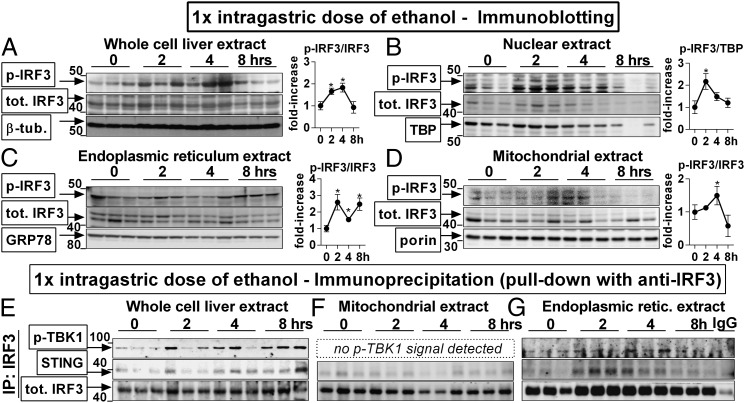
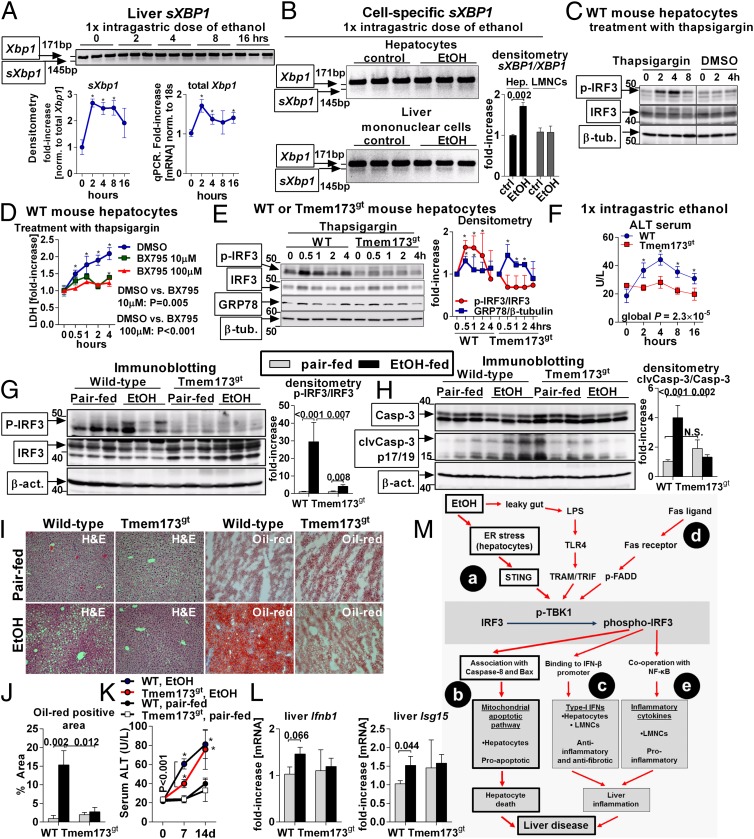
Emerging evidence suggests that innate immunity drives alcoholic liver disease (ALD) and that the interferon regulatory factor 3 (IRF3),a transcription factor regulating innate immune responses, is indispensable for the development of ALD. Here we report that IRF3 mediates ALD via linking endoplasmic reticulum (ER) stress with apoptotic signaling in hepatocytes. We found that ethanol induced ER stress and triggered the association of IRF3 with the ER adaptor, stimulator of interferon genes (STING), as well as subsequent phosphorylation of IRF3. Activated IRF3 associated with the proapoptotic molecule Bax [B-cell lymphoma 2 (Bcl2)-associated X protein] and contributed to hepatocyte apoptosis. Deficiency of STING prevented IRF3 phosphorylation by ethanol or ER stress, and absence of IRF3 prevented hepatocyte apoptosis. The pathogenic role of IRF3 in ALD was independent of inflammation or Type-I interferons. Thus, STING and IRF3 are key determinants of ALD, linking ER stress signaling with the mitochondrial pathway of hepatocyte apoptosis.
Keywords: Kupffer cells; interferon regulatory factor 3; steatohepatitis; stimulator of interferon genes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kumar H, Kawai T, Akira S. Pathogen recognition by the innate immune system. Int Rev Immunol. 2011;30(1):16–34. - PubMed
-
- Uesugi T, Froh M, Arteel GE, Bradford BU, Thurman RG. Toll-like receptor 4 is involved in the mechanism of early alcohol-induced liver injury in mice. Hepatology. 2001;34(1):101–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
