

Crystal structures of the CPAP/STIL complex reveal its role in centriole assembly and human microcephaly
- PMID: 24052813
- PMCID: PMC3776556
- DOI: 10.7554/eLife.01071
Crystal structures of the CPAP/STIL complex reveal its role in centriole assembly and human microcephaly
Abstract
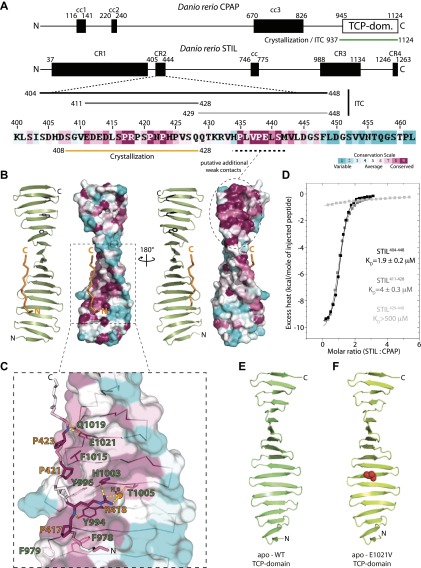
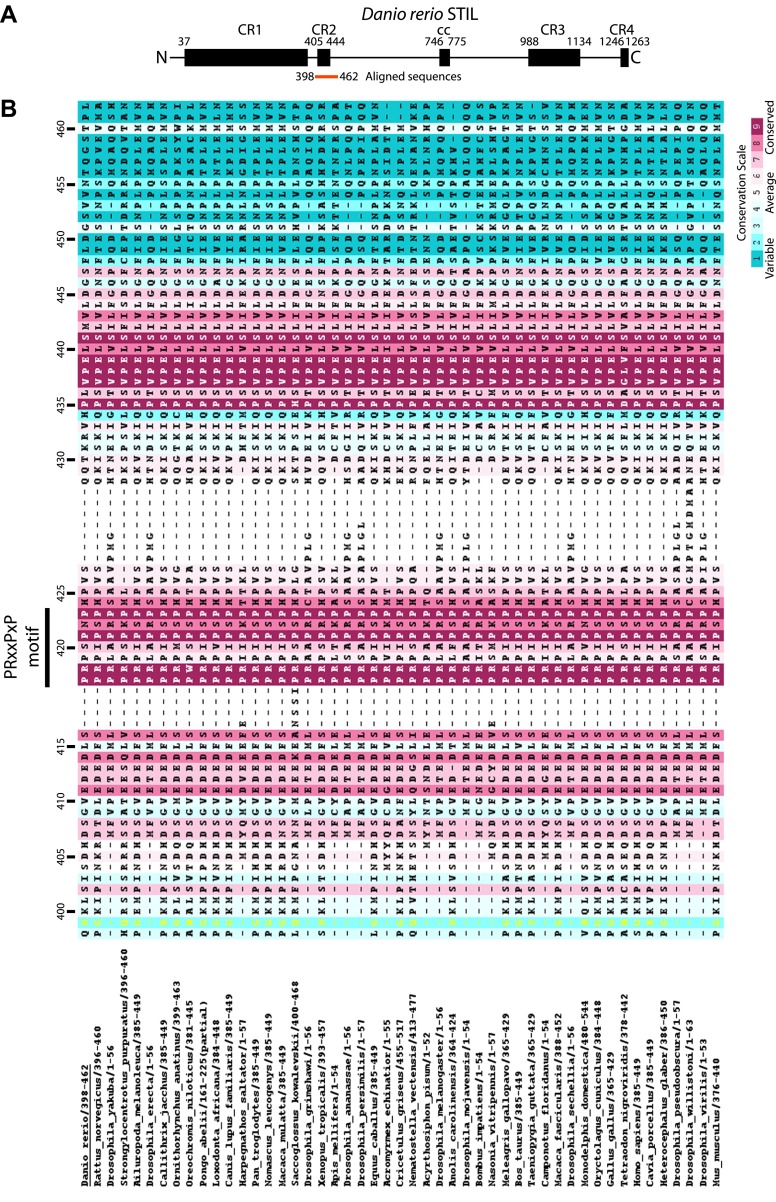
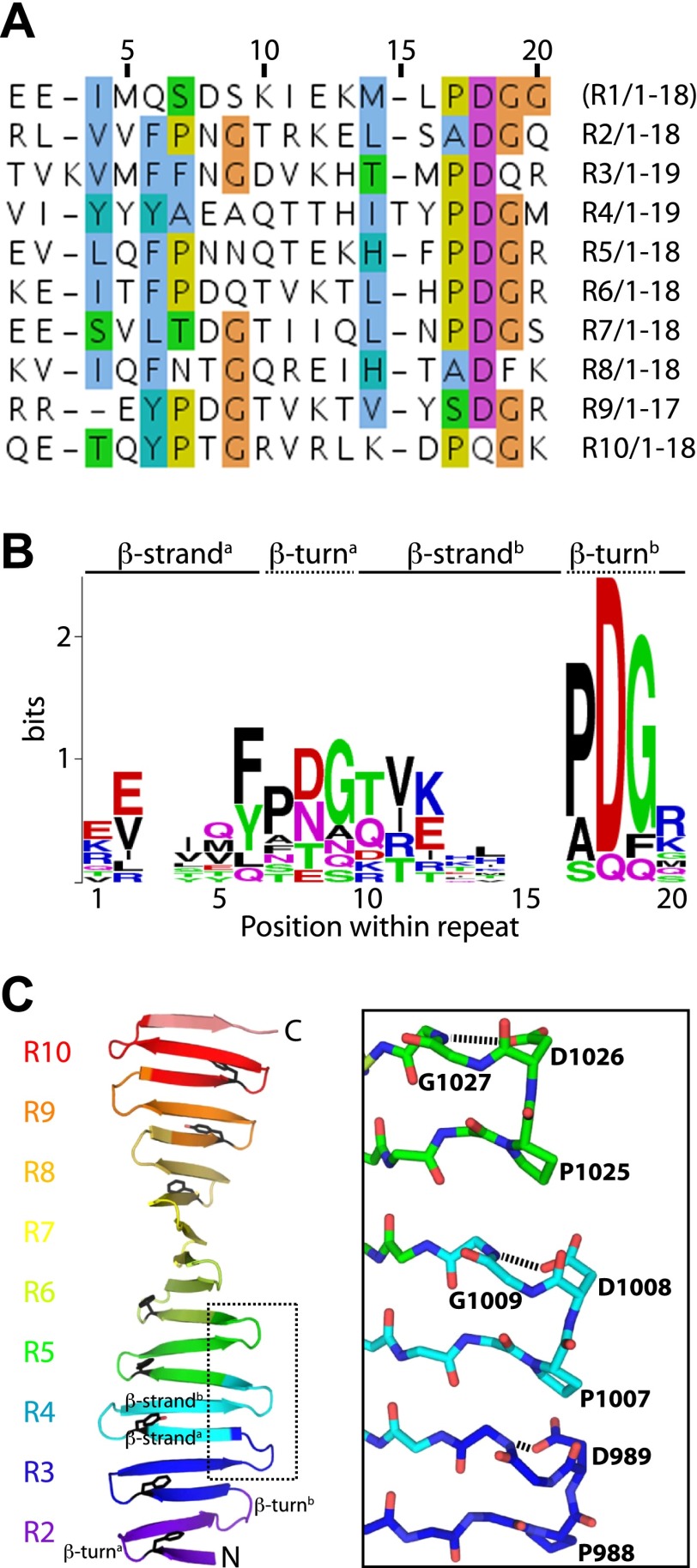
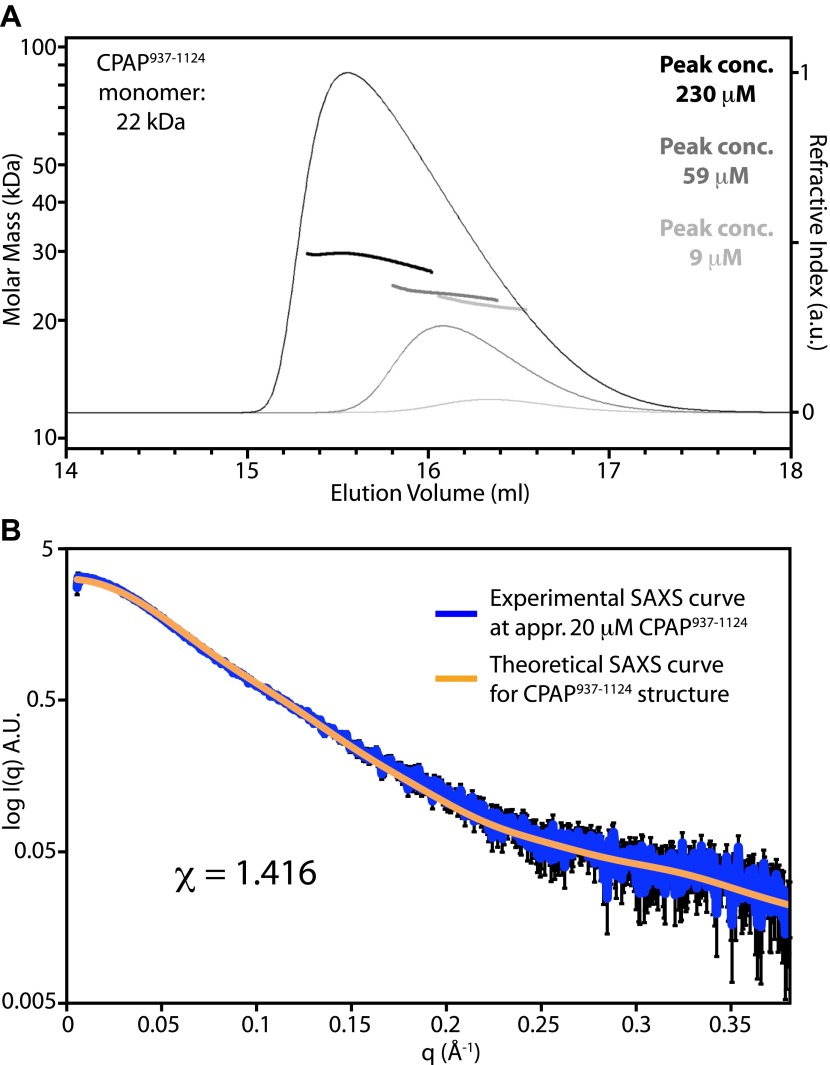
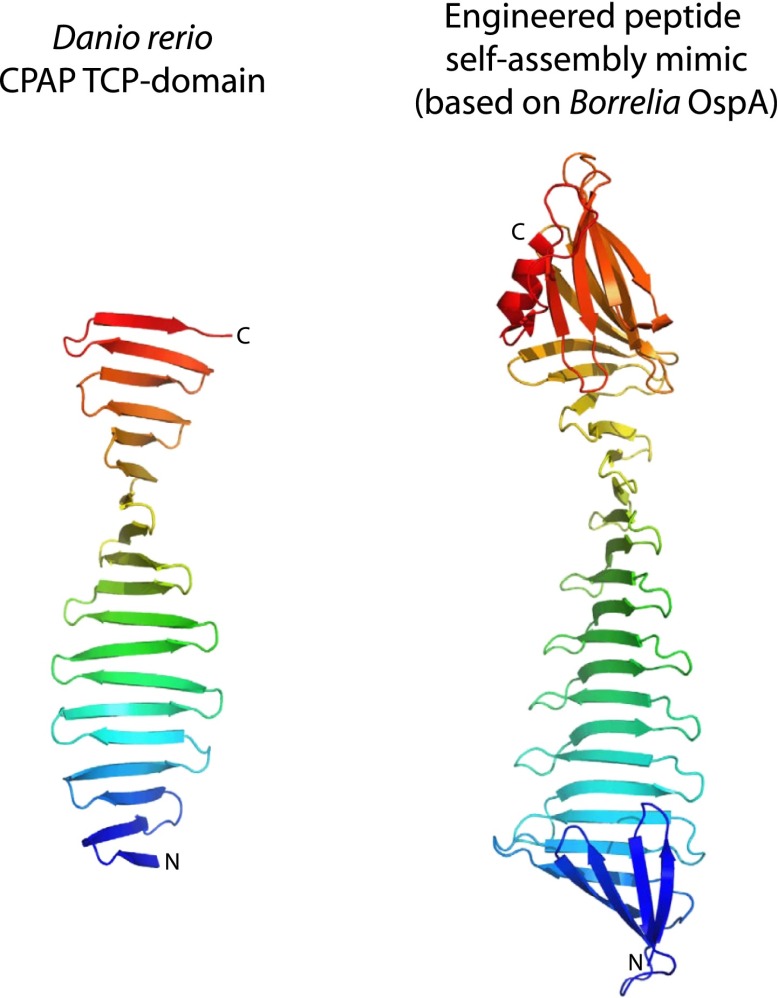
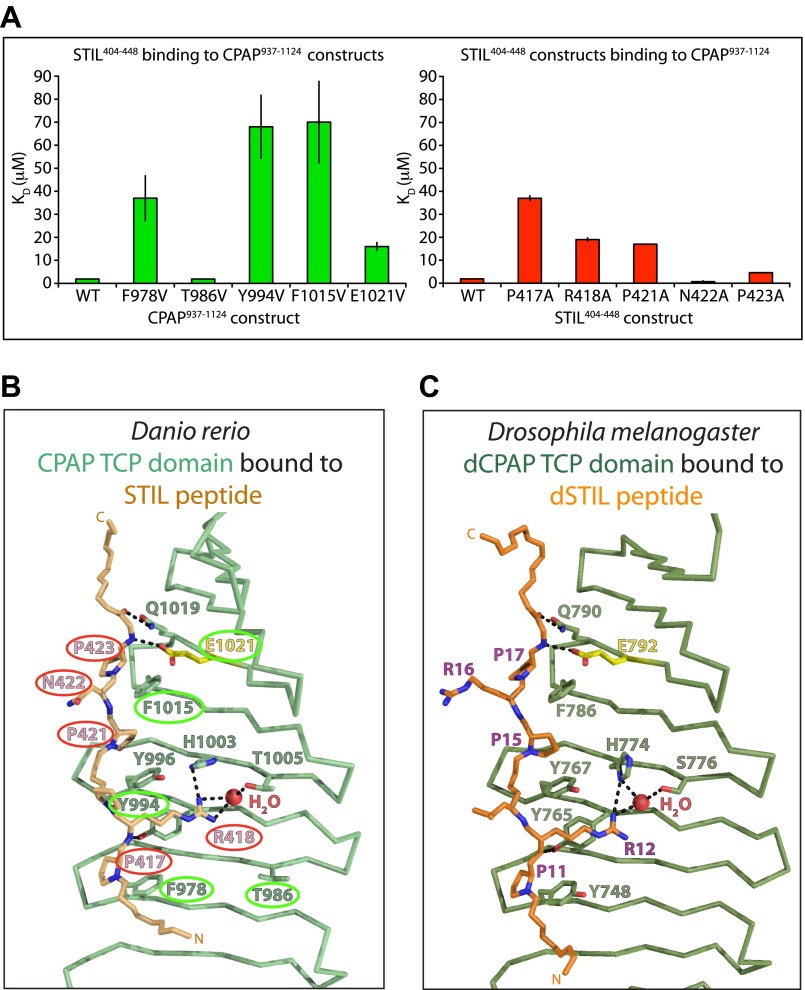
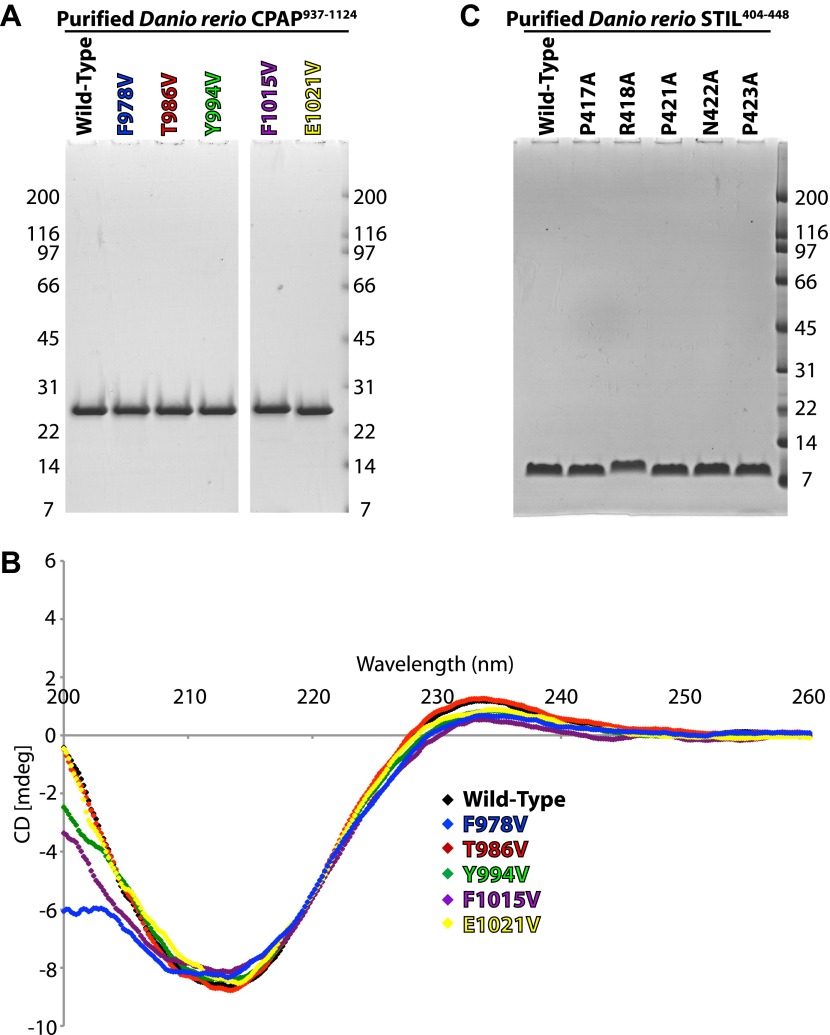
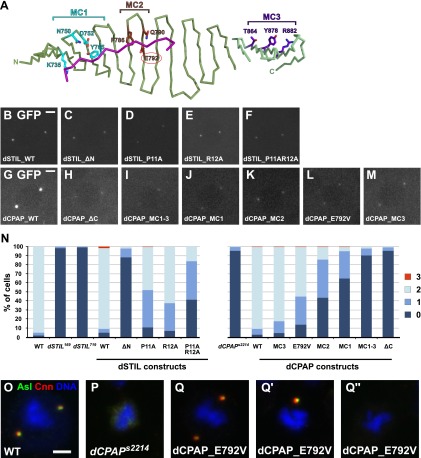
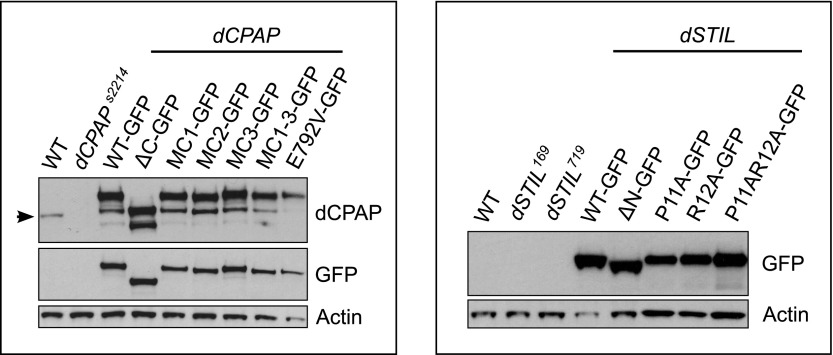
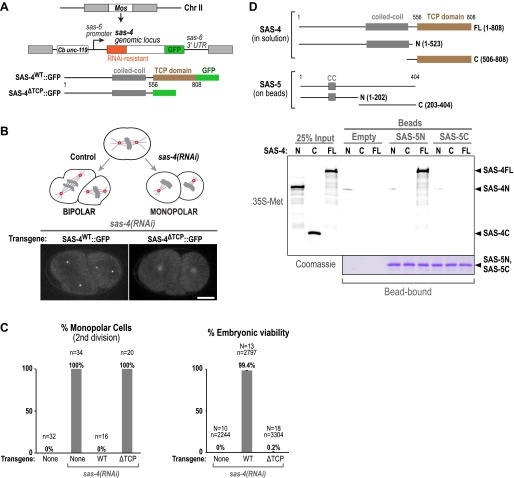
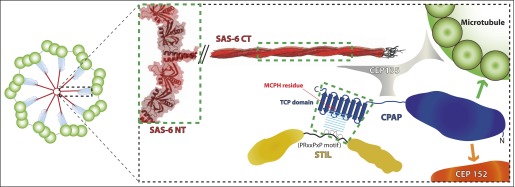
Centrioles organise centrosomes and template cilia and flagella. Several centriole and centrosome proteins have been linked to microcephaly (MCPH), a neuro-developmental disease associated with small brain size. CPAP (MCPH6) and STIL (MCPH7) are required for centriole assembly, but it is unclear how mutations in them lead to microcephaly. We show that the TCP domain of CPAP constitutes a novel proline recognition domain that forms a 1:1 complex with a short, highly conserved target motif in STIL. Crystal structures of this complex reveal an unusual, all-β structure adopted by the TCP domain and explain how a microcephaly mutation in CPAP compromises complex formation. Through point mutations, we demonstrate that complex formation is essential for centriole duplication in vivo. Our studies provide the first structural insight into how the malfunction of centriole proteins results in human disease and also reveal that the CPAP-STIL interaction constitutes a conserved key step in centriole biogenesis. DOI:http://dx.doi.org/10.7554/eLife.01071.001.
Keywords: C. elegans; CPAP; D. melanogaster; STIL; Zebrafish; centriole; centrosome; microcephaly.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
-
- Afonine PV, Grosse-Kunstleve RW, Adams PD. 2005. The Phenix refinement framework. CCP4 Newsletter 42 http://www.ccp4.ac.uk/newsletters/newsletter42/content.html
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
