

Membrane-sculpting BAR domains generate stable lipid microdomains
- PMID: 24055060
- PMCID: PMC4105227
- DOI: 10.1016/j.celrep.2013.08.024
Membrane-sculpting BAR domains generate stable lipid microdomains
Abstract
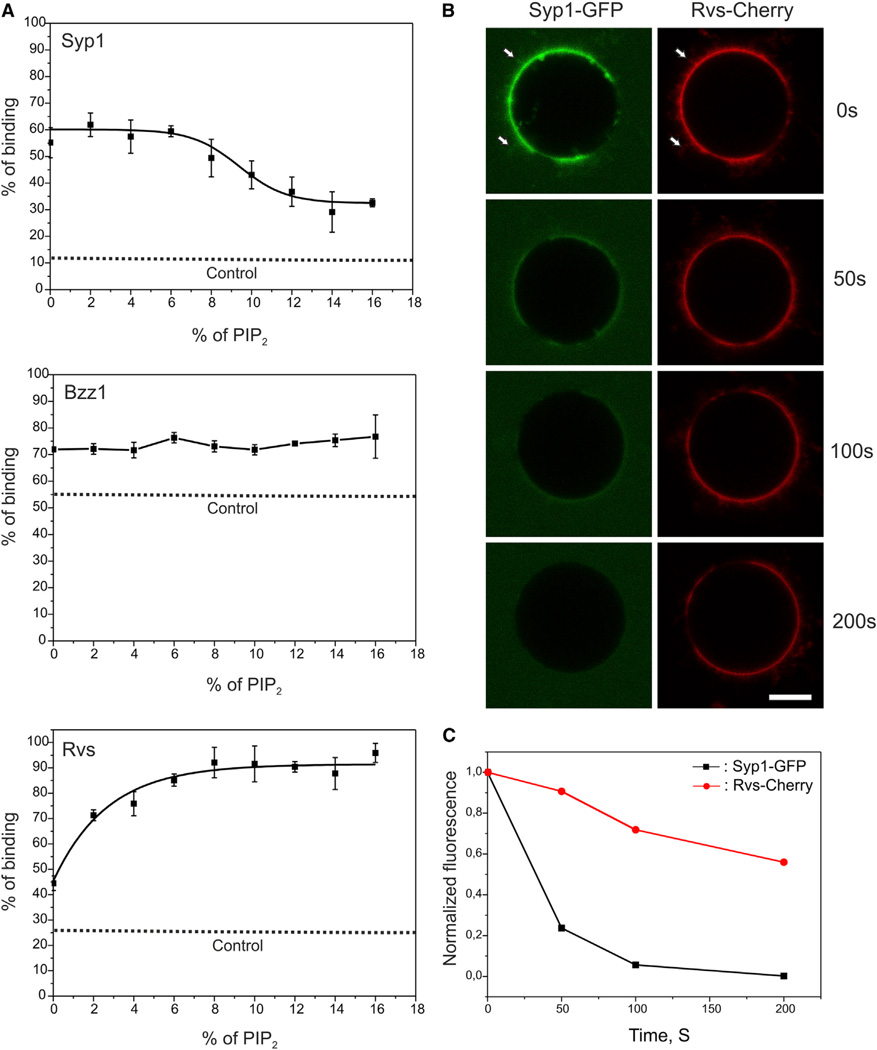
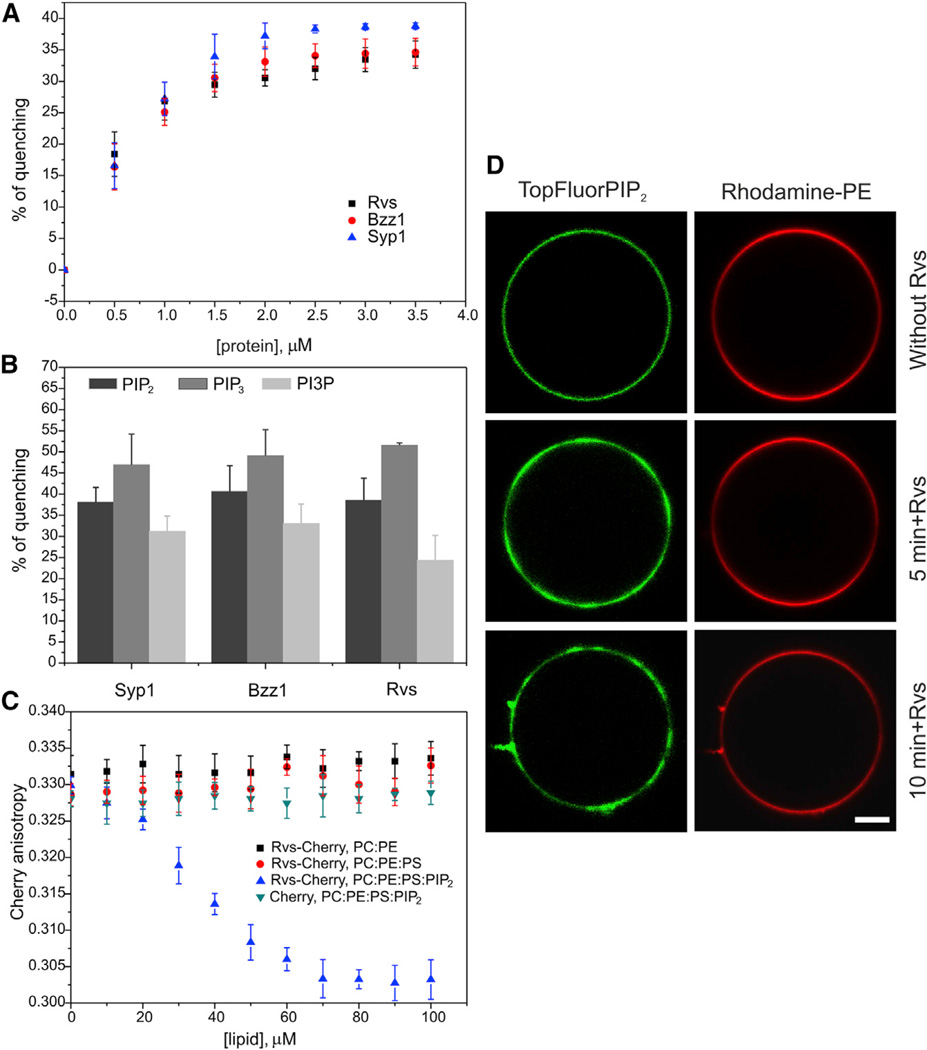
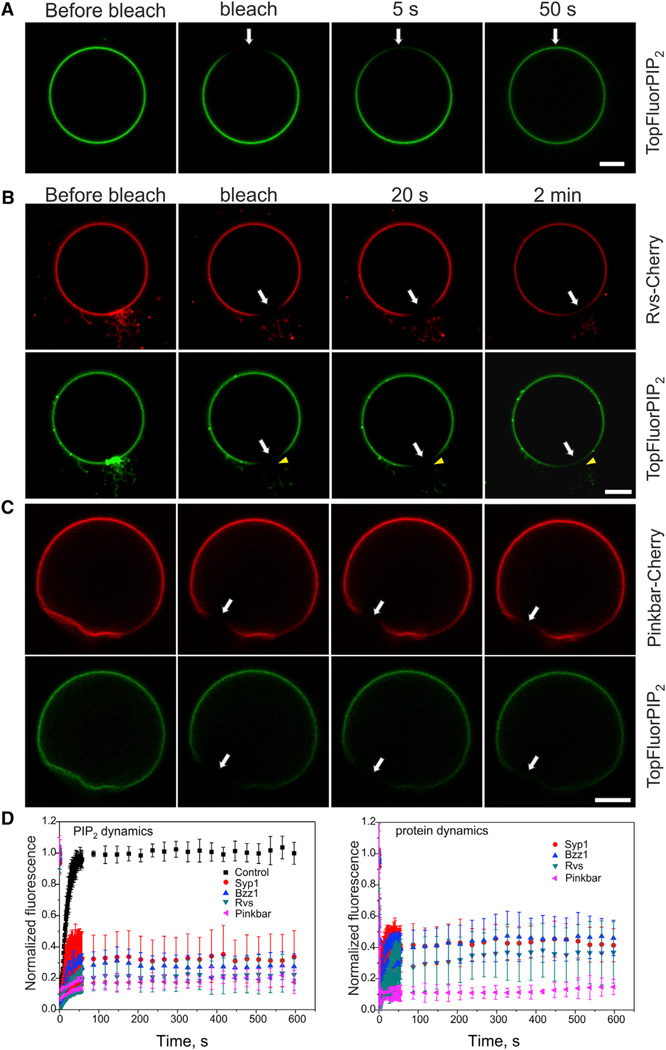
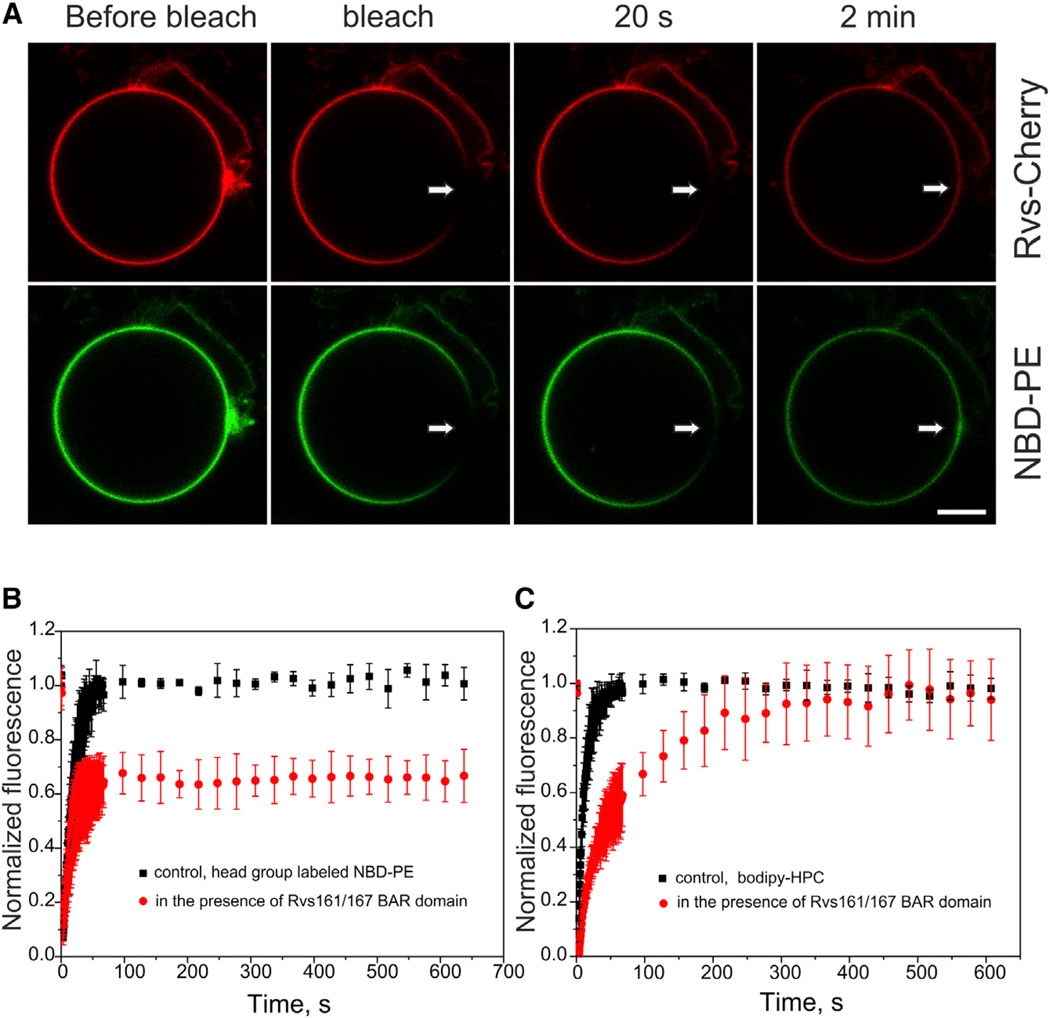
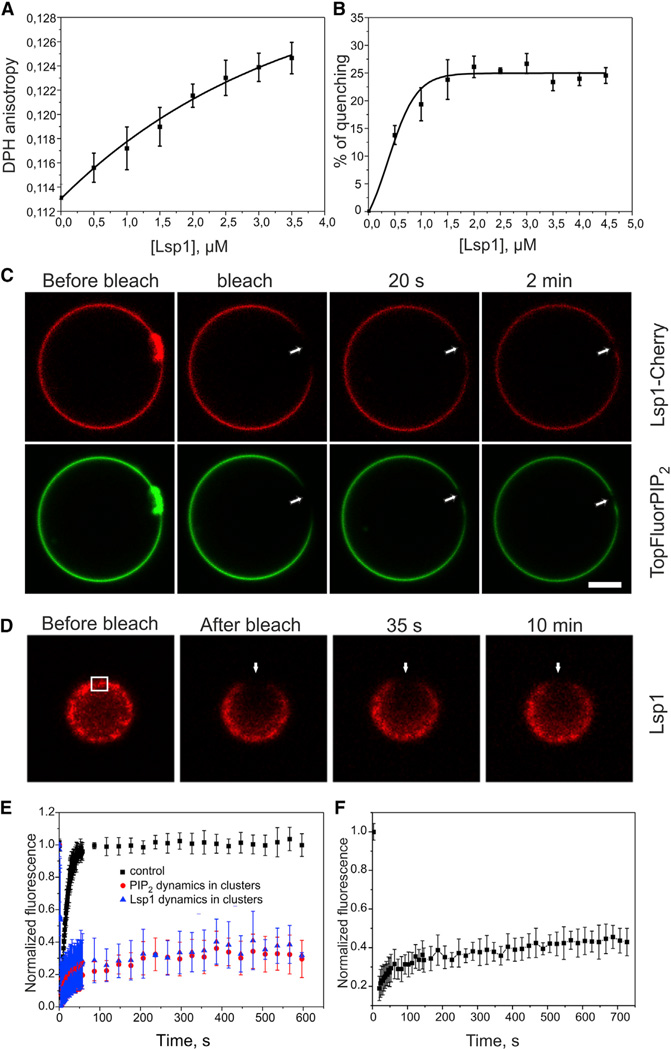
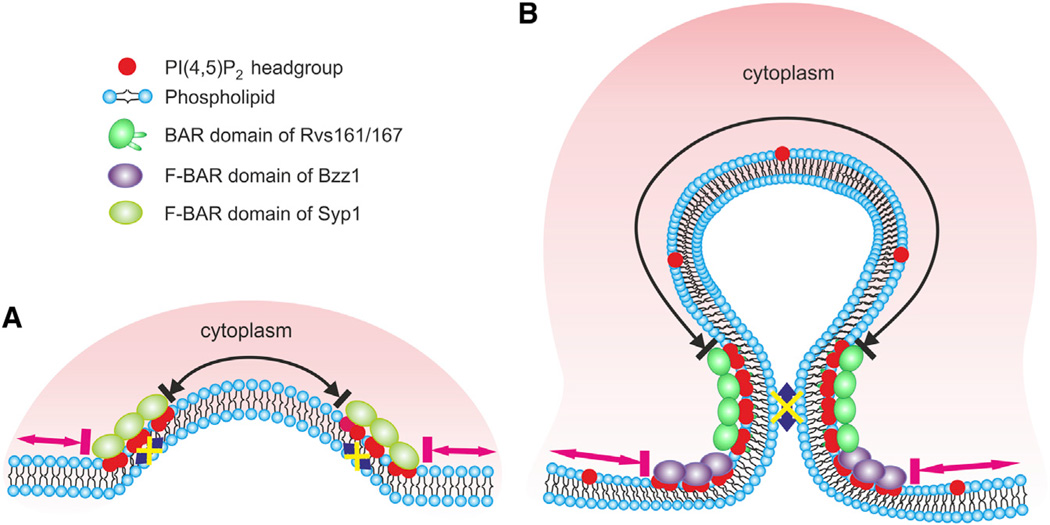
Bin-Amphiphysin-Rvs (BAR) domain proteins are central regulators of many cellular processes involving membrane dynamics. BAR domains sculpt phosphoinositide-rich membranes to generate membrane protrusions or invaginations. Here, we report that, in addition to regulating membrane geometry, BAR domains can generate extremely stable lipid microdomains by "freezing" phosphoinositide dynamics. This is a general feature of BAR domains, because the yeast endocytic BAR and Fes/CIP4 homology BAR (F-BAR) domains, the inverse BAR domain of Pinkbar, and the eisosomal BAR protein Lsp1 induced phosphoinositide clustering and halted lipid diffusion, despite differences in mechanisms of membrane interactions. Lsp1 displays comparable low diffusion rates in vitro and in vivo, suggesting that BAR domain proteins also generate stable phosphoinositide microdomains in cells. These results uncover a conserved role for BAR superfamily proteins in regulating lipid dynamics within membranes. Stable microdomains induced by BAR domain scaffolds and specific lipids can generate phase boundaries and diffusion barriers, which may have profound impacts on diverse cellular processes.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
