

A propofol binding site on mammalian GABAA receptors identified by photolabeling
- PMID: 24056400
- PMCID: PMC3951778
- DOI: 10.1038/nchembio.1340
A propofol binding site on mammalian GABAA receptors identified by photolabeling
Abstract
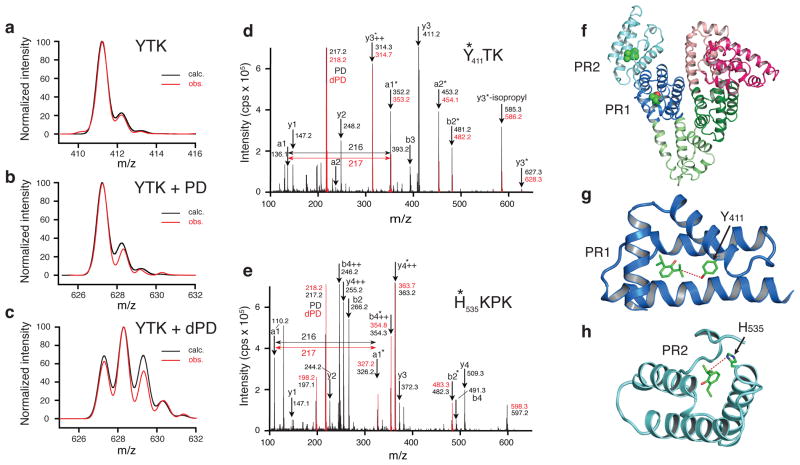
Propofol is the most important intravenous general anesthetic in current clinical use. It acts by potentiating GABAA (γ-aminobutyric acid type A) receptors, but where it binds to this receptor is not known and has been a matter of some debate. We synthesized a new propofol analog photolabeling reagent whose biological activity is very similar to that of propofol. We confirmed that this reagent labeled known propofol binding sites in human serum albumin that have been identified using X-ray crystallography. Using a combination of protiated and deuterated versions of the reagent to label mammalian receptors in intact membranes, we identified a new binding site for propofol in GABAA receptors consisting of both β3 homopentamers and α1β3 heteropentamers. The binding site is located within the β subunit at the interface between the transmembrane domains and the extracellular domain and lies close to known determinants of anesthetic sensitivity in the transmembrane segments TM1 and TM2.
Conflict of interest statement
Figures


References
-
- Jurd R, et al. General anesthetic actions in vivo strongly attenuated by a point mutation in the GABA(A) receptor beta3 subunit. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2003;17:250–252. - PubMed
-
- Zezula J, Slany A, Sieghart W. Interaction of allosteric ligands with GABAA receptors containing one, two, or three different subunits. European journal of pharmacology. 1996;301:207–214. - PubMed
-
- Feng HJ, Macdonald RL. Multiple actions of propofol on alphabetagamma and alphabetadelta GABAA receptors. Molecular pharmacology. 2004;66:1517–1524. - PubMed
Publication types
MeSH terms
Substances
Associated data
- PubChem-Substance/163835817
- PubChem-Substance/163835818
- PubChem-Substance/163835819
- PubChem-Substance/163835820
- PubChem-Substance/163835821
- PubChem-Substance/163835822
- PubChem-Substance/163835823
- PubChem-Substance/163835824
- PubChem-Substance/163835825
- PubChem-Substance/163835826
- PubChem-Substance/163835827
- PubChem-Substance/163835828
- PubChem-Substance/163835829
- PubChem-Substance/163835830
- PubChem-Substance/163835831
- PubChem-Substance/163835832
- PubChem-Substance/163835833
- PubChem-Substance/163835834
- PubChem-Substance/163835835
- PubChem-Substance/163835836
- PubChem-Substance/163835837
- PubChem-Substance/163835838
- PubChem-Substance/163835839
- PubChem-Substance/163835840
- PubChem-Substance/163835841
- PubChem-Substance/163835842
- PubChem-Substance/163835843
- PubChem-Substance/163835844
- PubChem-Substance/163835845
- PubChem-Substance/163835846
- PubChem-Substance/163835847
- PubChem-Substance/163835848
- PubChem-Substance/163835849
