

Direct regulation of cytochrome c oxidase by calcium ions
- PMID: 24058566
- PMCID: PMC3769247
- DOI: 10.1371/journal.pone.0074436
Direct regulation of cytochrome c oxidase by calcium ions
Abstract
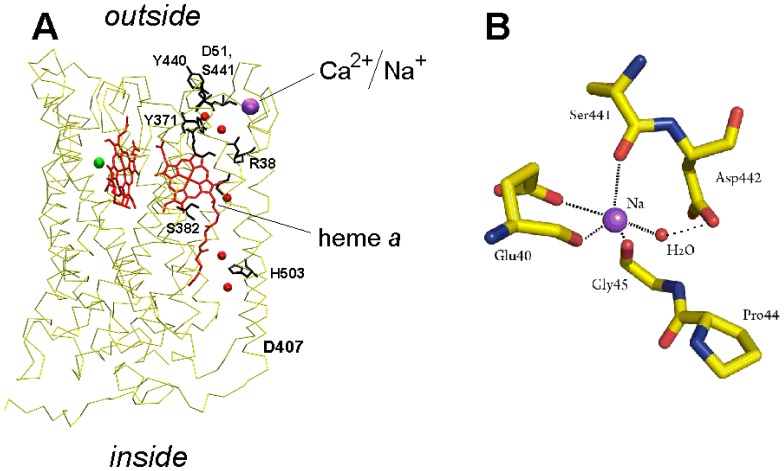
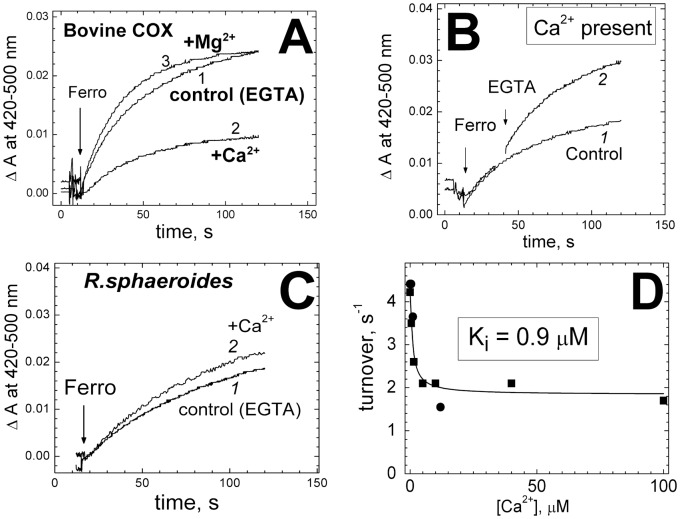
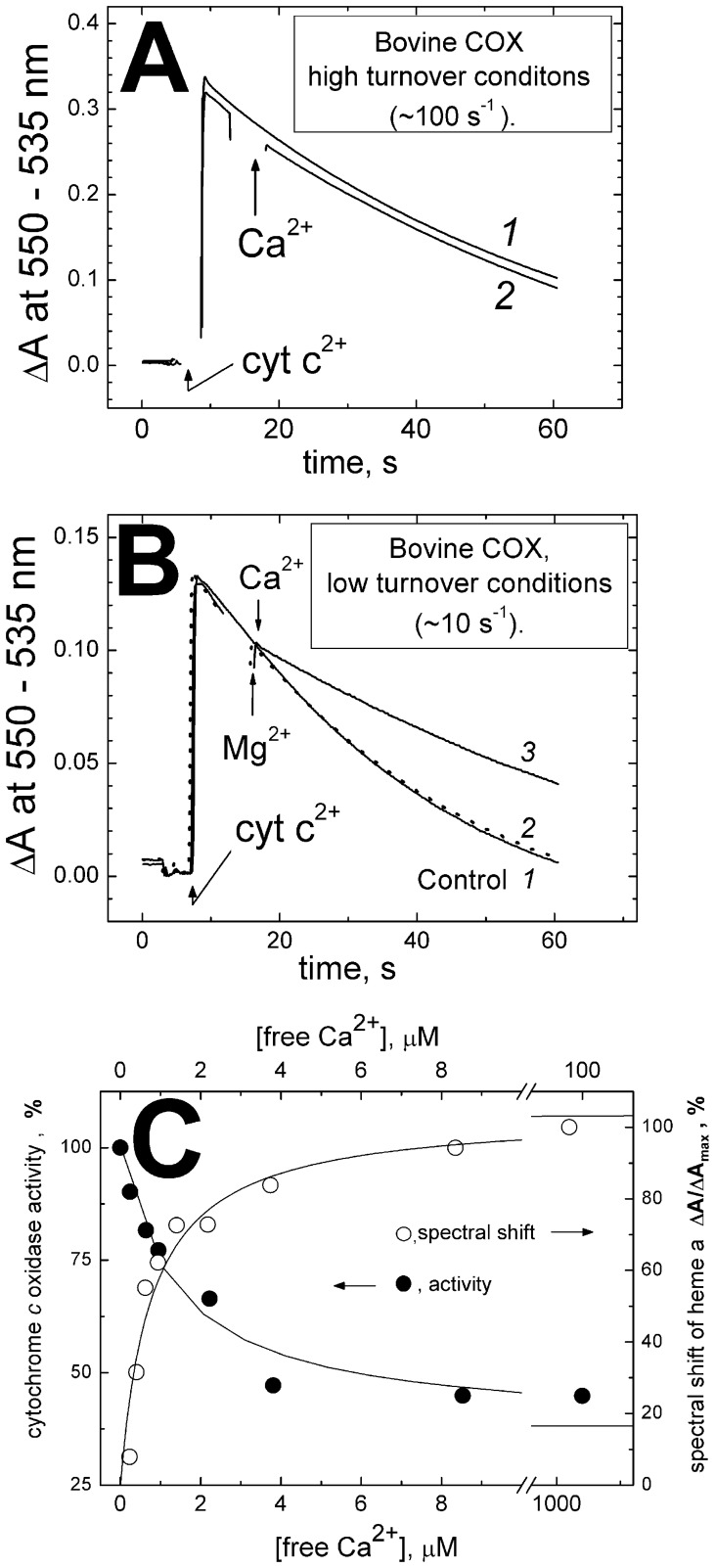
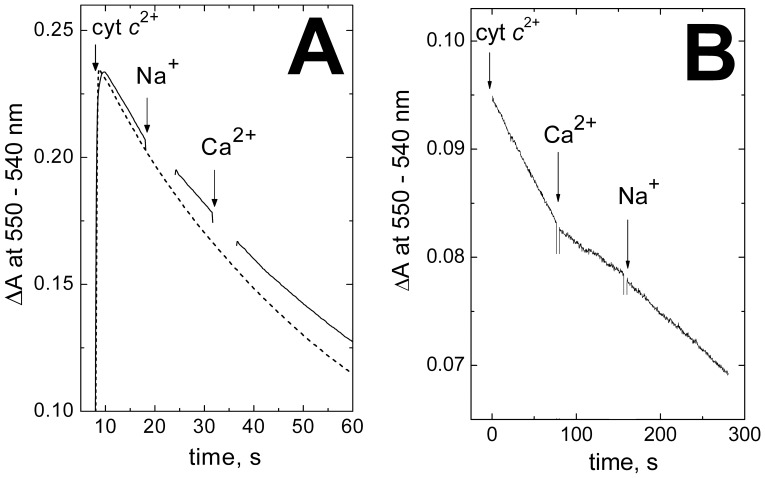
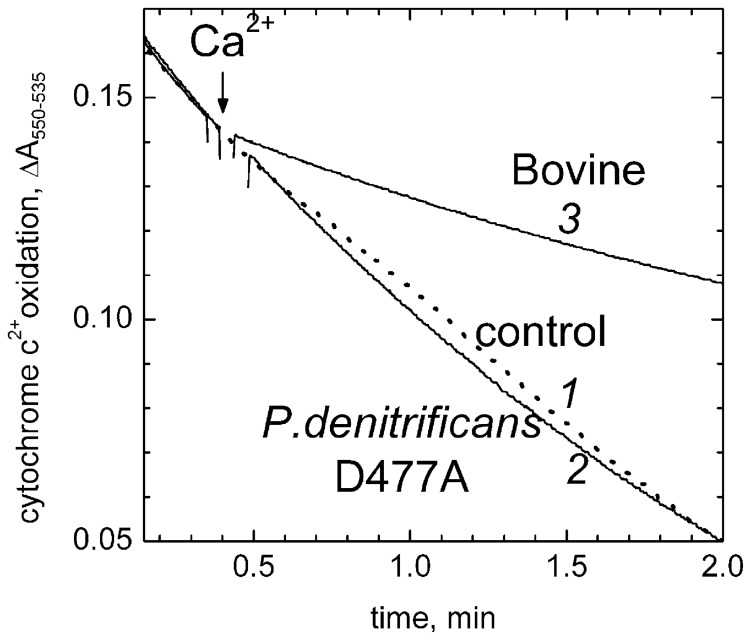
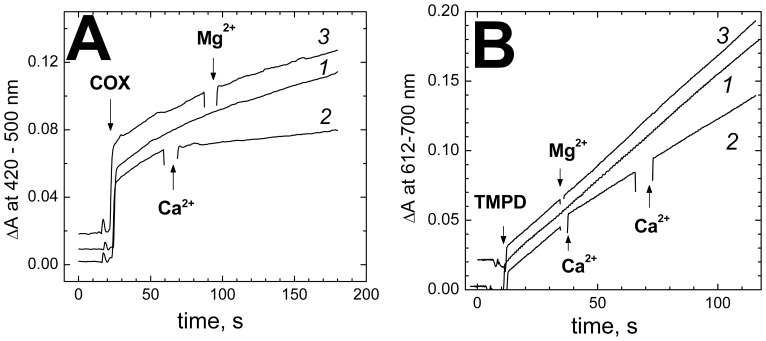
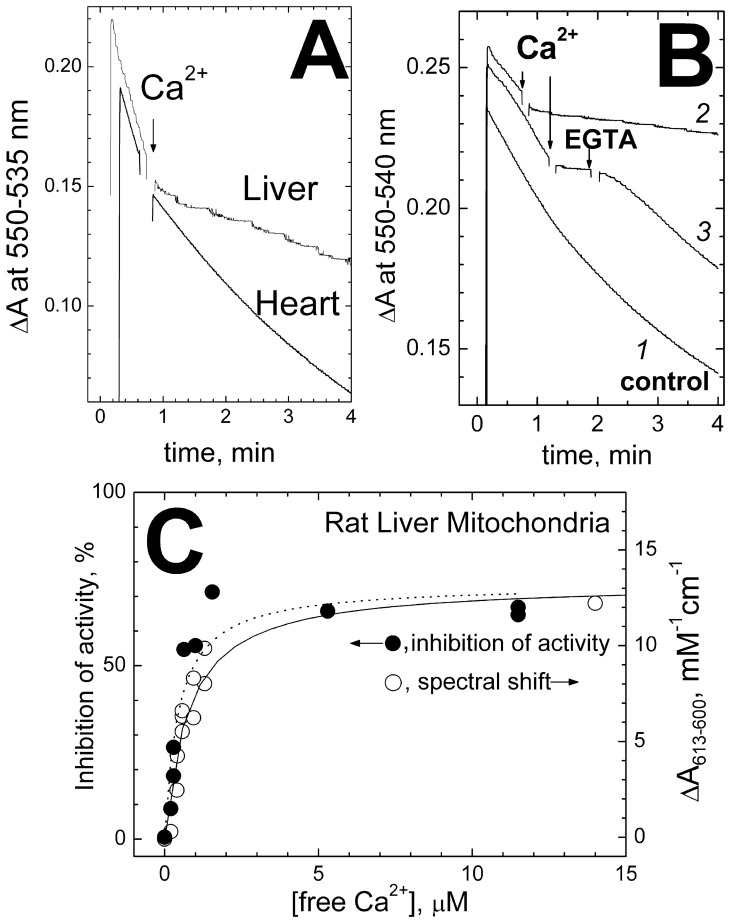
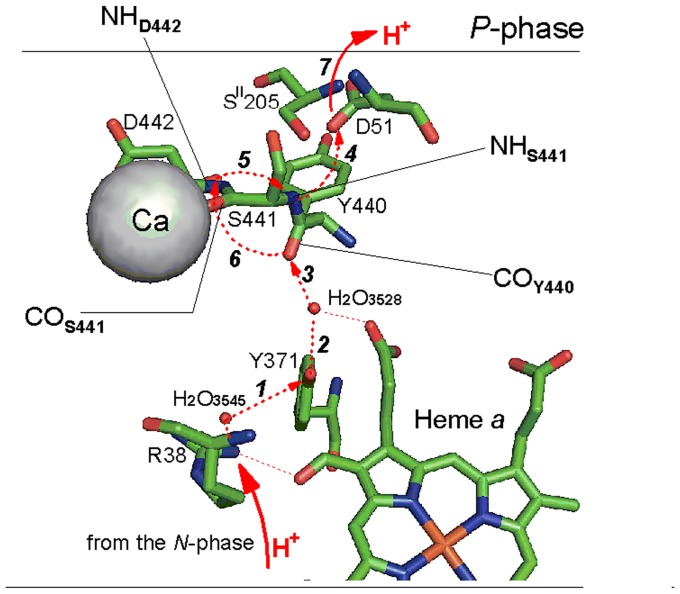
Cytochrome c oxidase from bovine heart binds Ca(2+) reversibly at a specific Cation Binding Site located near the outer face of the mitochondrial membrane. Ca(2+) shifts the absorption spectrum of heme a, which allowed previously to determine the kinetics and equilibrium characteristics of the binding. However, no effect of Ca(2+) on the functional characteristics of cytochrome oxidase was revealed earlier. Here we report that Ca(2+) inhibits cytochrome oxidase activity of isolated bovine heart enzyme by 50-60% with Ki of ∼1 µM, close to Kd of calcium binding with the oxidase determined spectrophotometrically. The inhibition is observed only at low, but physiologically relevant, turnover rates of the enzyme (∼10 s(-1) or less). No inhibitory effect of Ca(2+) is observed under conventional conditions of cytochrome c oxidase activity assays (turnover number >100 s(-1) at pH 8), which may explain why the effect was not noticed earlier. The inhibition is specific for Ca(2+) and is reversed by EGTA. Na(+) ions that compete with Ca(2+) for binding with the Cation Binding Site, do not affect significantly activity of the enzyme but counteract the inhibitory effect of Ca(2+). The Ca(2+)-induced inhibition of cytochrome c oxidase is observed also with the uncoupled mitochondria from several rat tissues. At the same time, calcium ions do not inhibit activity of the homologous bacterial cytochrome oxidases. Possible mechanisms of the inhibition are discussed as well as potential physiological role of Ca(2+) binding with cytochrome oxidase. Ca(2+)- binding at the Cation Binding Site is proposed to inhibit proton-transfer through the exit part of the proton conducting pathway H in the mammalian oxidases.
Conflict of interest statement
Figures
References
-
- Ferguson-Miller S, Babcock GT (1996) Heme/Copper Terminal Oxidases. Chem Rev 7: 2889–2907. - PubMed
-
- Belevich I, Verkhovsky MI (2008) Molecular Mechanism of Proton Translocation by Cytochrome c Oxidase. Antioxidants and Redox Signaling 10: 1–29. - PubMed
-
- Einarsdottir O, Caughey W (1985) Bovine heart cytochrome c oxidase preparations contain high affinity binding sites for magnesium as well as for zinc, copper, and heme iron. Biochem Biophys Res Commun 129: 840–847. - PubMed
-
- Iwata S, Ostermeier C, Ludwig B, Michel H (1995) Structure at 2.8 A Resolution of Cytochrome c Oxidase from Paracoccus denitrificans . Nature 376: 660–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
