

Point mutations in the transmembrane region of the clic1 ion channel selectively modify its biophysical properties
- PMID: 24058583
- PMCID: PMC3776819
- DOI: 10.1371/journal.pone.0074523
Point mutations in the transmembrane region of the clic1 ion channel selectively modify its biophysical properties
Abstract
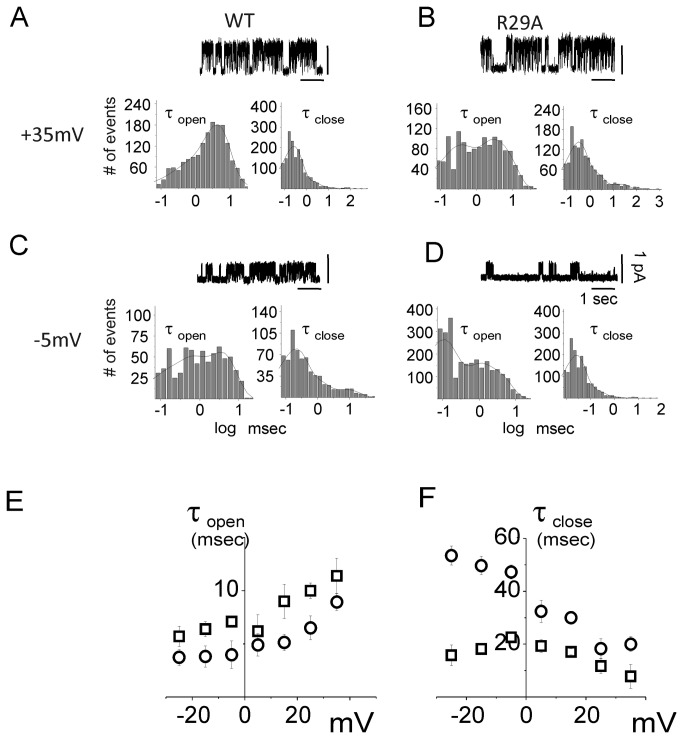
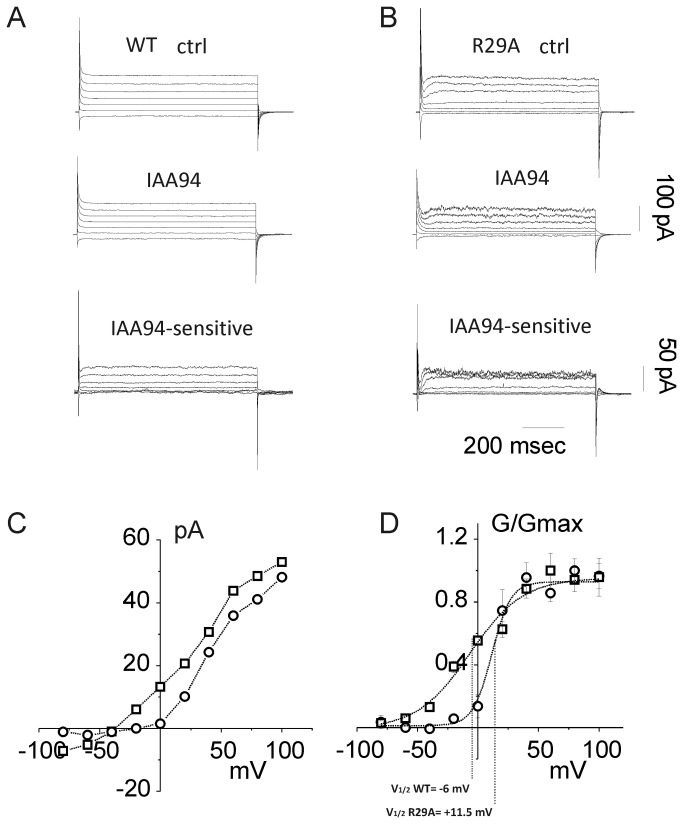
Chloride intracellular Channel 1 (CLIC1) is a metamorphic protein that changes from a soluble cytoplasmic protein into a transmembrane protein. Once inserted into membranes, CLIC1 multimerises and is able to form chloride selective ion channels. Whilst CLIC1 behaves as an ion channel both in cells and in artificial lipid bilayers, its structure in the soluble form has led to some uncertainty as to whether it really is an ion channel protein. CLIC1 has a single putative transmembrane region that contains only two charged residues: arginine 29 (Arg29) and lysine 37 (Lys37). As charged residues are likely to have a key role in ion channel function, we hypothesized that mutating them to neutral alanine to generate K37A and R29A CLIC1 would alter the electrophysiological characteristics of CLIC1. By using three different electrophysiological approaches: i) single channel Tip-Dip in artificial bilayers using soluble recombinant CLIC1, ii) cell-attached and iii) whole-cell patch clamp recordings in transiently transfected HEK cells, we determined that the K37A mutation altered the single-channel conductance while the R29A mutation affected the single-channel open probability in response to variation in membrane potential. Our results show that mutation of the two charged amino acids (K37 and R29) in the putative transmembrane region of CLIC1 alters the biophysical properties of the ion channel in both artificial bilayers and cells. Hence these charged residues are directly involved in regulating its ion channel activity. This strongly suggests that, despite its unusual structure, CLIC1 itself is able to form a chloride ion channel.
Conflict of interest statement
Figures

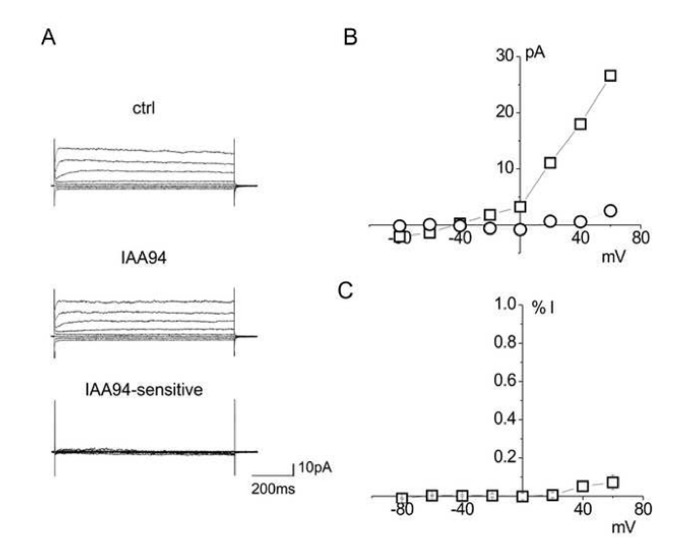
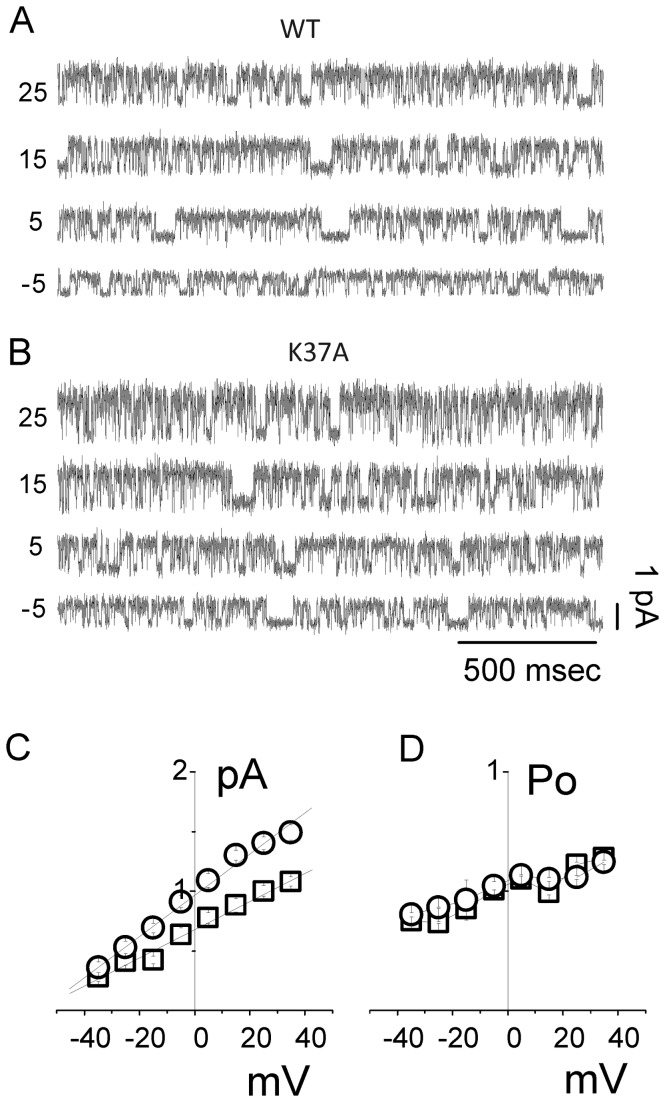
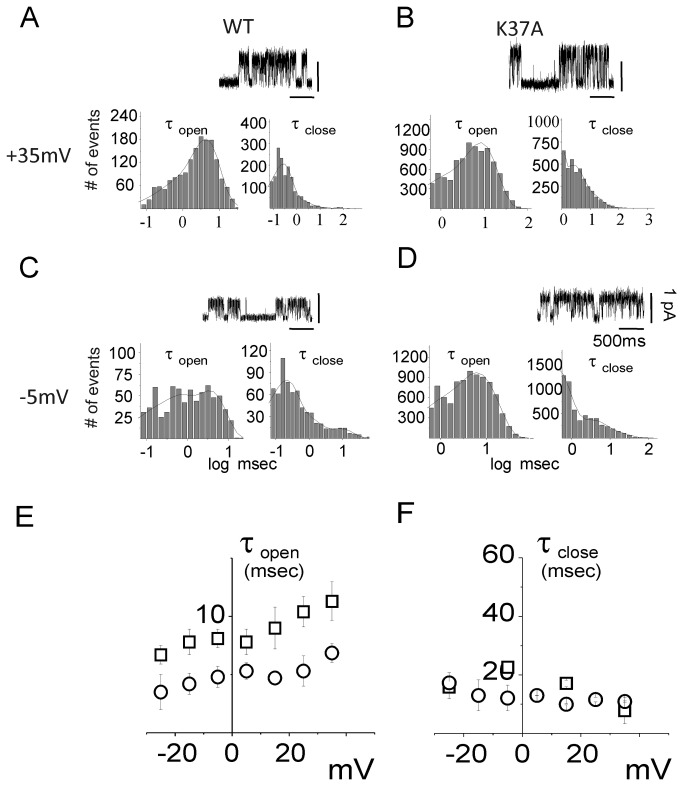
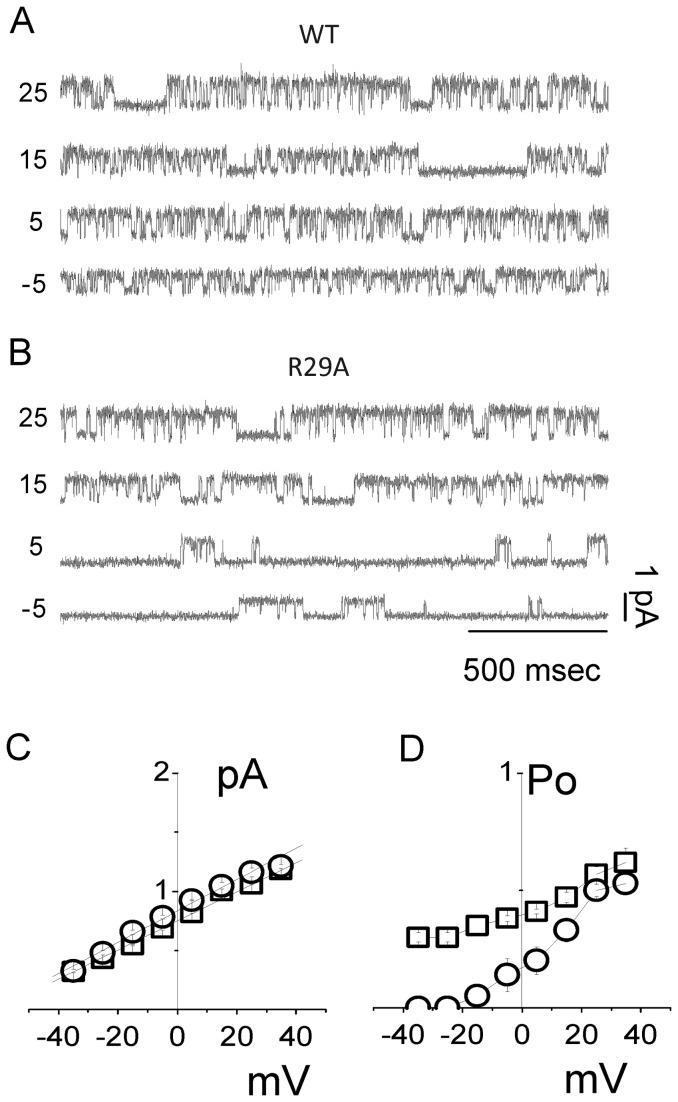
References
-
- Valenzuela SM, Martin DK, Por SB, Robbins JM, Warton K et al. (1997) Molecular cloning and expression of a chloride ion channel of cell nuclei. J Biol Chem 272: 12575-12582. doi:10.1074/jbc.272.19.12575. PubMed: 9139710. - DOI - PubMed
-
- Yang SG, Wang WY, Ling TJ, Feng Y, Du XT et al. (2010) alpha-Tocopherol quinone inhibits beta-amyloid aggregation and cytotoxicity, disaggregates preformed fibrils and decreases the production of reactive oxygen species, NO and inflammatory cytokines. Neurochem Int 57: 914-922. doi:10.1016/j.neuint.2010.09.011. PubMed: 20933033. - DOI - PubMed
-
- Littler DR, Harrop SJ, Goodchild SC, Phang JM, Mynott AV et al. (2010) The enigma of the CLIC proteins: Ion channels, redox proteins, enzymes, scaffolding proteins? FEBS Lett 584: 2093-2101. doi:10.1016/j.febslet.2010.01.027. PubMed: 20085760. - DOI - PubMed
-
- Averaimo S, Milton RH, Duchen MR, Mazzanti M (2010) Chloride intracellular channel 1 (CLIC1): Sensor and effector during oxidative stress. FEBS Lett 584: 2076-2084. doi:10.1016/j.febslet.2010.02.073. PubMed: 20385134. - DOI - PubMed
-
- Edwards JC, Kahl CR (2010) Chloride channels of intracellular membranes. FEBS Lett 584: 2102-2111. doi:10.1016/j.febslet.2010.01.037. PubMed: 20100480. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
