

Chromium(VI) causes interstrand DNA cross-linking in vitro but shows no hypersensitivity in cross-link repair-deficient human cells
- PMID: 24059640
- PMCID: PMC3838998
- DOI: 10.1021/tx400293s
Chromium(VI) causes interstrand DNA cross-linking in vitro but shows no hypersensitivity in cross-link repair-deficient human cells
Abstract
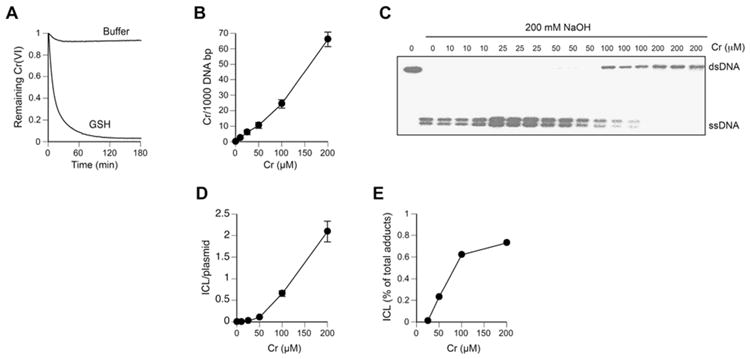
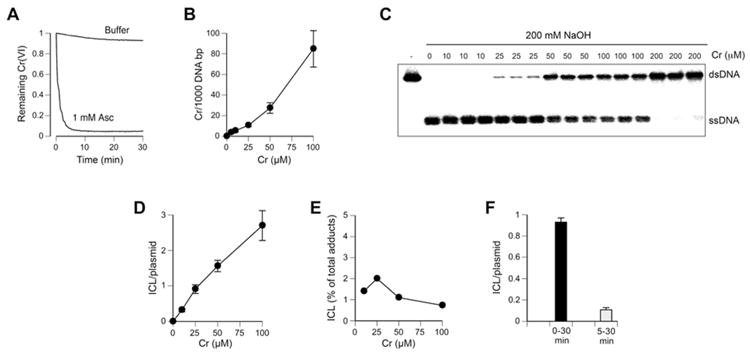
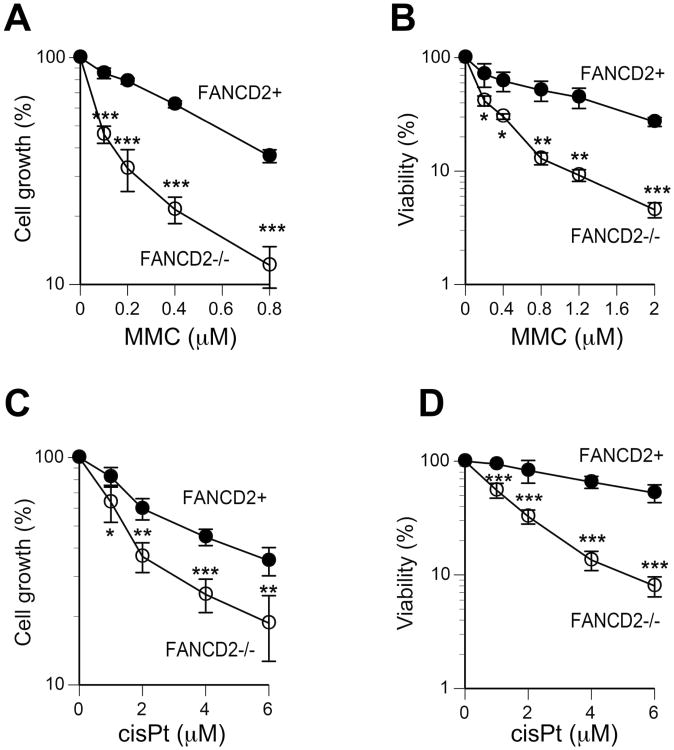
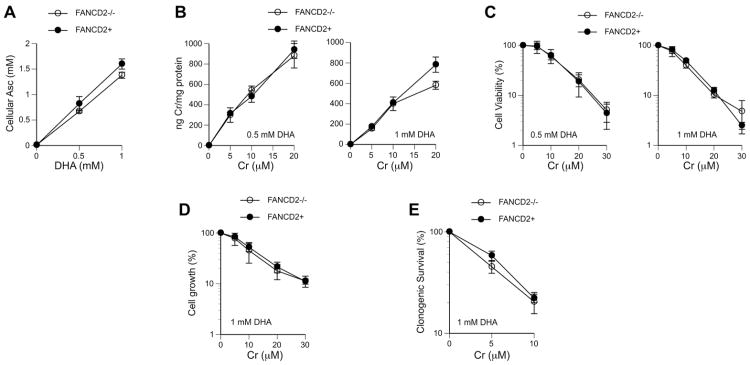
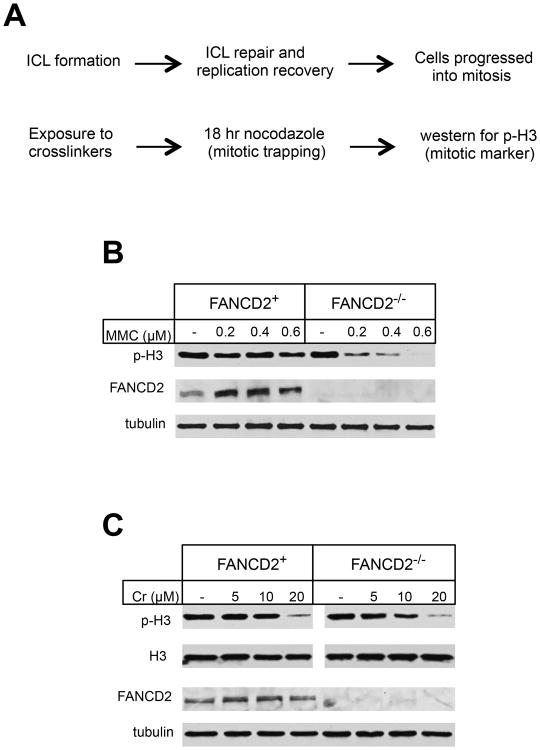
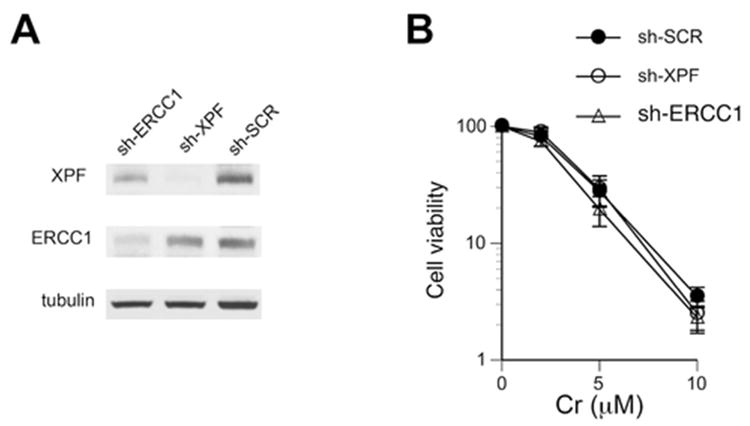
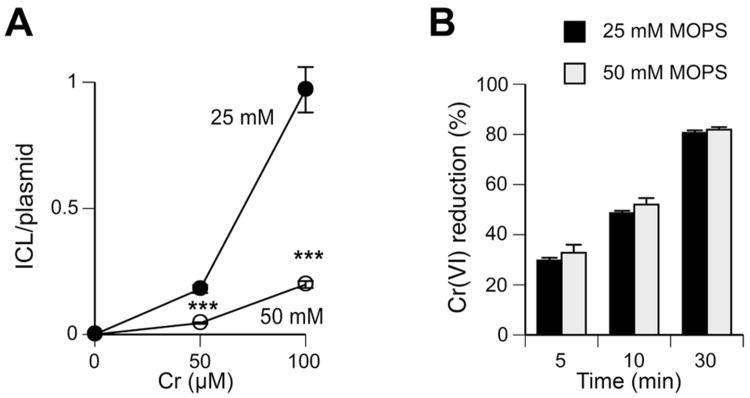
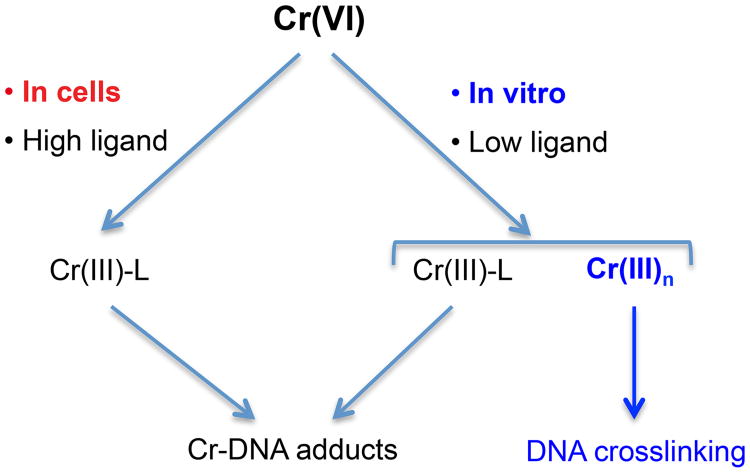
Hexavalent chromium is a human carcinogen activated primarily by direct reduction with cellular ascorbate and to a lesser extent, by glutathione. Cr(III), the final product of Cr(VI) reduction, forms six bonds allowing intermolecular cross-linking. In this work, we investigated the ability of Cr(VI) to cause interstrand DNA cross-links (ICLs) whose formation mechanisms and presence in human cells are currently uncertain. We found that in vitro reduction of Cr(VI) with glutathione showed a sublinear production of ICLs, the yield of which was less than 1% of total Cr-DNA adducts at the optimal conditions. Formation of ICLs in fast ascorbate-Cr(VI) reactions occurred during a short reduction interval and displayed a linear dose dependence with the average yield of 1.3% of total adducts. In vitro production of ICLs was strongly suppressed by increasing buffer molarity, indicating inhibitory effects of ligand-Cr(III) binding on the formation of cross-linking species. The presence of ICLs in human cells was assessed from the impact of ICL repair deficiencies on Cr(VI) responses. We found that ascorbate-restored FANCD2-null and isogenic FANCD2-complemented cells showed similar cell cycle inhibition and toxicity by Cr(VI). XPA-null cells are defective in the repair of Cr-DNA monoadducts, but stable knockdowns of ERCC1 or XPF in these cells with extended time for the completion of cross-linking reactions did not produce any sensitization to Cr(VI). Our results together with chemical and steric considerations of Cr(III) reactivity suggest that ICL generation by chromate is probably an in vitro phenomenon occurring at conditions permitting the formation of Cr(III) oligomers.
Figures
Similar articles
-
XPF-ERCC1 participates in the Fanconi anemia pathway of cross-link repair.Mol Cell Biol. 2009 Dec;29(24):6427-37. doi: 10.1128/MCB.00086-09. Epub 2009 Oct 5. Mol Cell Biol. 2009. PMID: 19805513 Free PMC article.
-
Critical role of chromium (Cr)-DNA interactions in the formation of Cr-induced polymerase arresting lesions.Biochemistry. 2002 Oct 15;41(41):12529-37. doi: 10.1021/bi020452j. Biochemistry. 2002. PMID: 12369844
-
Genotoxicity and mutagenicity of chromium(VI)/ascorbate-generated DNA adducts in human and bacterial cells.Biochemistry. 2003 Feb 4;42(4):1062-70. doi: 10.1021/bi0271547. Biochemistry. 2003. PMID: 12549927
-
Complexities of chromium carcinogenesis: role of cellular response, repair and recovery mechanisms.Mutat Res. 2003 Dec 10;533(1-2):3-36. doi: 10.1016/j.mrfmmm.2003.09.006. Mutat Res. 2003. PMID: 14643411 Review.
-
Chromium(VI) reduction by ascorbate: role of reactive intermediates in DNA damage in vitro.Environ Health Perspect. 1994 Sep;102 Suppl 3(Suppl 3):21-5. doi: 10.1289/ehp.94102s321. Environ Health Perspect. 1994. PMID: 7843100 Free PMC article. Review.
Cited by
-
Tdp1 processes chromate-induced single-strand DNA breaks that collapse replication forks.PLoS Genet. 2018 Aug 27;14(8):e1007595. doi: 10.1371/journal.pgen.1007595. eCollection 2018 Aug. PLoS Genet. 2018. PMID: 30148840 Free PMC article.
-
Homologous Recombination and Translesion DNA Synthesis Play Critical Roles on Tolerating DNA Damage Caused by Trace Levels of Hexavalent Chromium.PLoS One. 2016 Dec 1;11(12):e0167503. doi: 10.1371/journal.pone.0167503. eCollection 2016. PLoS One. 2016. PMID: 27907204 Free PMC article.
-
Chromium-catechin complex, synthesis and toxicity check using bacterial models.Heliyon. 2020 Aug 6;6(8):e04563. doi: 10.1016/j.heliyon.2020.e04563. eCollection 2020 Aug. Heliyon. 2020. PMID: 32793825 Free PMC article.
-
Forming a chromium-based interstrand DNA crosslink: Implications for carcinogenicity.J Inorg Biochem. 2024 Feb;251:112439. doi: 10.1016/j.jinorgbio.2023.112439. Epub 2023 Nov 25. J Inorg Biochem. 2024. PMID: 38039560 Free PMC article.
-
Formaldehyde-Assisted Isolation of Regulatory Elements (FAIRE) analysis uncovers broad changes in chromatin structure resulting from hexavalent chromium exposure.PLoS One. 2014 May 16;9(5):e97849. doi: 10.1371/journal.pone.0097849. eCollection 2014. PLoS One. 2014. PMID: 24837440 Free PMC article.
References
-
- Langard S. One hundred years of chromium and cancer: a review of epidemiological evidence and selected case reports. Am J Ind Med. 1990;17:189–215. - PubMed
-
- Costa M, Klein CB. Toxicity and carcinogenicity of chromium compounds in humans. Crit Rev Toxicol. 2006;36:155–163. - PubMed
-
- Suzuki Y, Fukuda K. Reduction of hexavalent chromium by ascorbic acid and glutathione with special reference to the rat lung. Arch Toxicol. 1990;64:169–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
