

HIV-1 envelope glycoprotein signatures that correlate with the development of cross-reactive neutralizing activity
- PMID: 24059682
- PMCID: PMC3849187
- DOI: 10.1186/1742-4690-10-102
HIV-1 envelope glycoprotein signatures that correlate with the development of cross-reactive neutralizing activity
Abstract
Background: Current HIV-1 envelope glycoprotein (Env) vaccines are unable to induce cross-reactive neutralizing antibodies. However, such antibodies are elicited in 10-30% of HIV-1 infected individuals, but it is unknown why these antibodies are induced in some individuals and not in others. We hypothesized that the Envs of early HIV-1 variants in individuals who develop cross-reactive neutralizing activity (CrNA) might have unique characteristics that support the induction of CrNA.
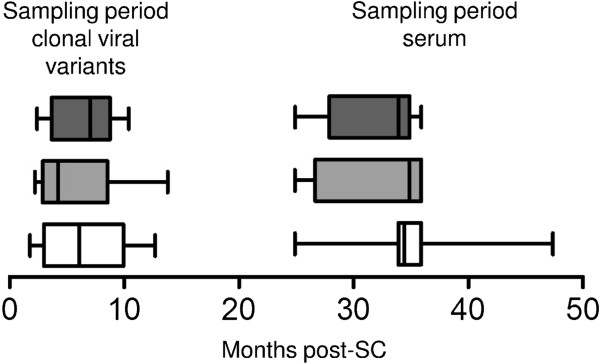
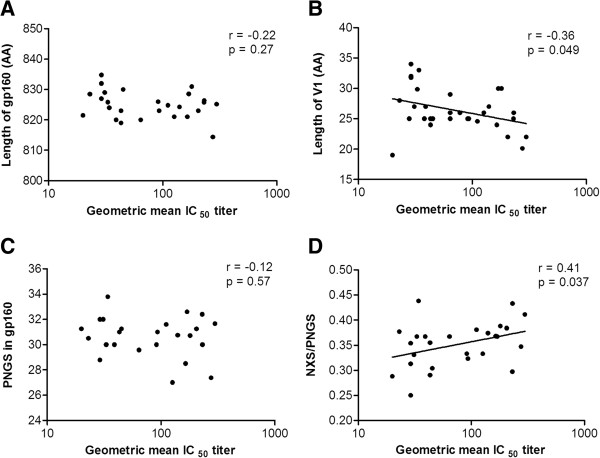
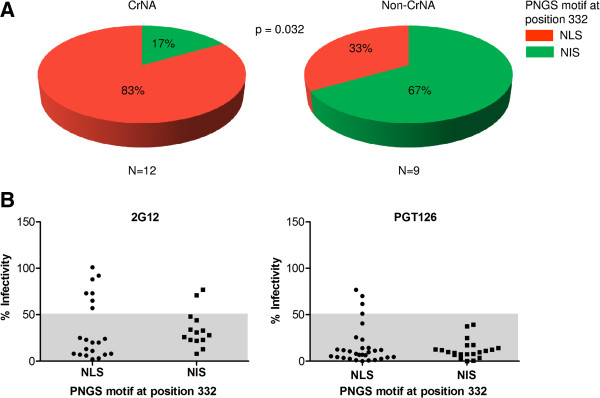
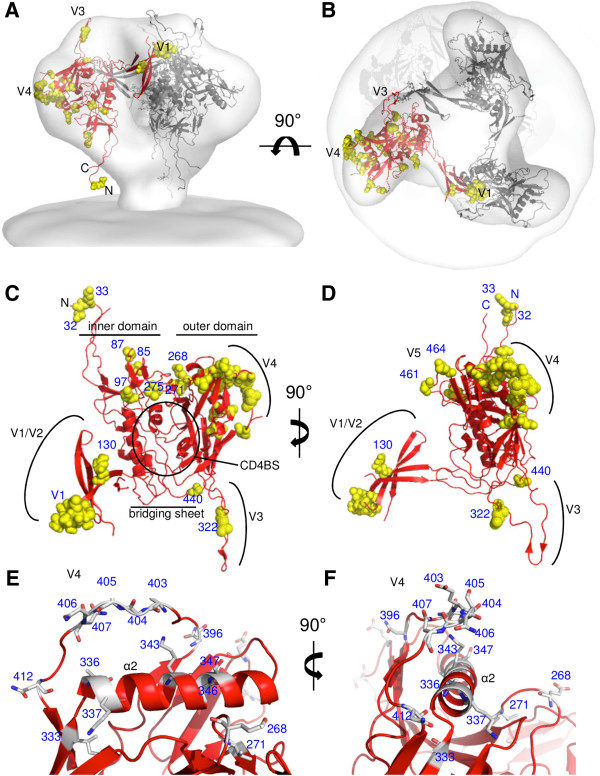
Results: We retrospectively generated and analyzed env sequences of early HIV-1 clonal variants from 31 individuals with diverse levels of CrNA 2-4 years post-seroconversion. These sequences revealed a number of Env signatures that coincided with CrNA development. These included a statistically shorter variable region 1 and a lower probability of glycosylation as implied by a high ratio of NXS versus NXT glycosylation motifs. Furthermore, lower probability of glycosylation at position 332, which is involved in the epitopes of many broadly reactive neutralizing antibodies, was associated with the induction of CrNA. Finally, Sequence Harmony identified a number of amino acid changes associated with the development of CrNA. These residues mapped to various Env subdomains, but in particular to the first and fourth variable region as well as the underlying α2 helix of the third constant region.
Conclusions: These findings imply that the development of CrNA might depend on specific characteristics of early Env. Env signatures that correlate with the induction of CrNA might be relevant for the design of effective HIV-1 vaccines.
Figures
References
-
- Belshe RB, Graham BS, Keefer MC, Gorse GJ, Wright P, Dolin R, Matthews T, Weinhold K, Bolognesi DP, Sposto R. et al. Neutralizing antibodies to HIV-1 in seronegative volunteers immunized with recombinant gp120 from the MN strain of HIV-1. NIAID AIDS Vaccine Clinical Trials Network. JAMA. 1994;272:475–480. - PubMed
-
- Mascola JR, Snyder SW, Weislow OS, Belay SM, Belshe RB, Schwartz DH, Clements ML, Dolin R, Graham BS, Gorse GJ. et al. Immunization with envelope subunit vaccine products elicits neutralizing antibodies against laboratory-adapted but not primary isolates of human immunodeficiency virus type 1. J Infect Dis. 1996;173:340–348. - PubMed
-
- Pitisuttithum P, Berman PW, Phonrat B, Suntharasamai P, Raktham S, Srisuwanvilai LO, Hirunras K, Kitayaporn D, Kaewkangwal J, Migasena S. et al. Phase I/II study of a candidate vaccine designed against the B and E subtypes of HIV-1. J Acquir Immune Defic Syndr. 2004;37:1160–1165. - PubMed
-
- Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E. et al. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand. N Engl J Med. 2009;361:2209–2220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
