

Divergent evolution of ligand binding in the o-succinylbenzoate synthase family
- PMID: 24060347
- PMCID: PMC3908897
- DOI: 10.1021/bi401176d
Divergent evolution of ligand binding in the o-succinylbenzoate synthase family
Abstract
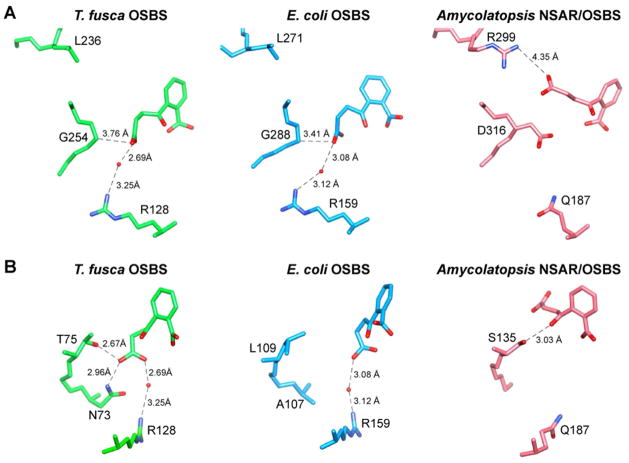
Thermobifida fusca o-succinylbenzoate synthase (OSBS), a member of the enolase superfamily that catalyzes a step in menaquinone biosynthesis, has an amino acid sequence that is 22 and 28% identical with those of two previously characterized OSBS enzymes from Escherichia coli and Amycolatopsis sp. T-1-60, respectively. These values are considerably lower than typical levels of sequence identity among homologous proteins that have the same function. To determine how such divergent enzymes catalyze the same reaction, we determined the structure of T. fusca OSBS and identified amino acids that are important for ligand binding. We discovered significant differences in structure and conformational flexibility between T. fusca OSBS and other members of the enolase superfamily. In particular, the 20s loop, a flexible loop in the active site that permits ligand binding and release in most enolase superfamily proteins, has a four-amino acid deletion and is well-ordered in T. fusca OSBS. Instead, the flexibility of a different region allows the substrate to enter from the other side of the active site. T. fusca OSBS was more tolerant of mutations at residues that were critical for activity in E. coli OSBS. Also, replacing active site amino acids found in one protein with the amino acids that occur at the same place in the other protein reduces the catalytic efficiency. Thus, the extraordinary divergence between these proteins does not appear to reflect a higher tolerance of mutations. Instead, large deletions outside the active site were accompanied by alteration of active site size and electrostatic interactions, resulting in small but significant differences in ligand binding.
Figures




References
-
- Gerlt JA, Babbitt PC, Rayment I. Divergent evolution in the enolase superfamily: the interplay of mechanism and specificity. Arch Biochem Biophys. 2005;433:59–70. - PubMed
-
- Rakus JF, Kalyanaraman C, Fedorov AA, Fedorov EV, Mills-Groninger FP, Toro R, Bonanno J, Bain K, Sauder JM, Burley SK, Almo SC, Jacobson MP, Gerlt JA. Computation-facilitated assignment of the function in the enolase superfamily: a regiochemically distinct galactarate dehydratase from Oceanobacillus iheyensis. Biochemistry. 2009;48:11546–11558. - PMC - PubMed
-
- Song L, Kalyanaraman C, Fedorov AA, Fedorov EV, Glasner ME, Brown S, Imker HJ, Babbitt PC, Almo SC, Jacobson MP, Gerlt JA. Prediction and assignment of function for a divergent N-succinyl amino acid racemase. Nat Chem Biol. 2007;3:486–491. - PubMed
-
- Lukk T, Sakai A, Kalyanaraman C, Brown SD, Imker HJ, Song L, Fedorov AA, Fedorov EV, Toro R, Hillerich B, Seidel R, Patskovsky Y, Vetting MW, Nair SK, Babbitt PC, Almo SC, Gerlt JA, Jacobson MP. Homology models guide discovery of diverse enzyme specificities among dipeptide epimerases in the enolase superfamily. Proc Natl Acad Sci U S A. 2012;109:4122–4127. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
