

Compositionality in neural control: an interdisciplinary study of scribbling movements in primates
- PMID: 24062679
- PMCID: PMC3771313
- DOI: 10.3389/fncom.2013.00103
Compositionality in neural control: an interdisciplinary study of scribbling movements in primates
Abstract
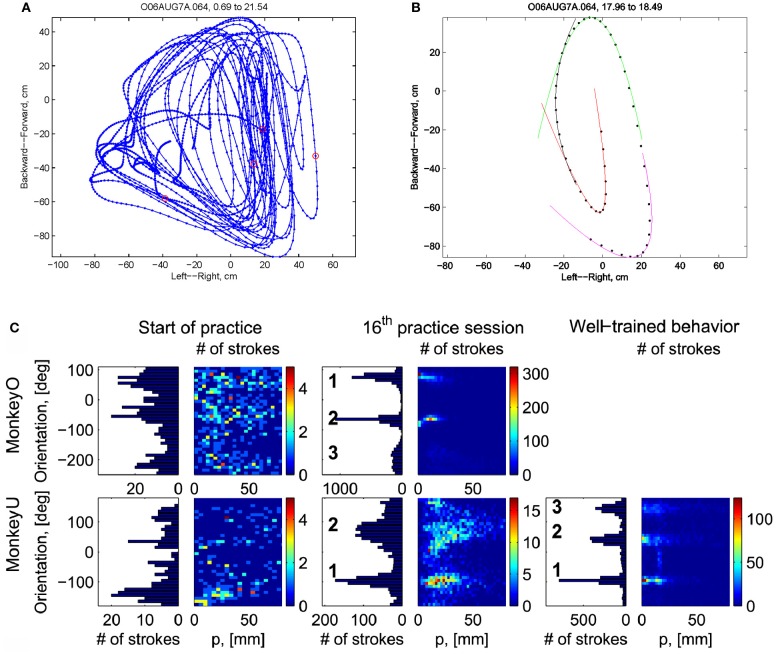
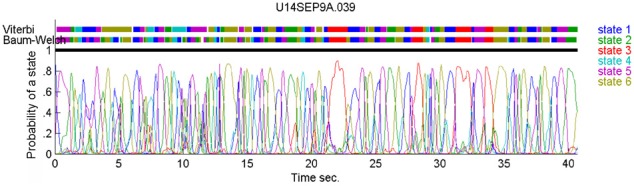
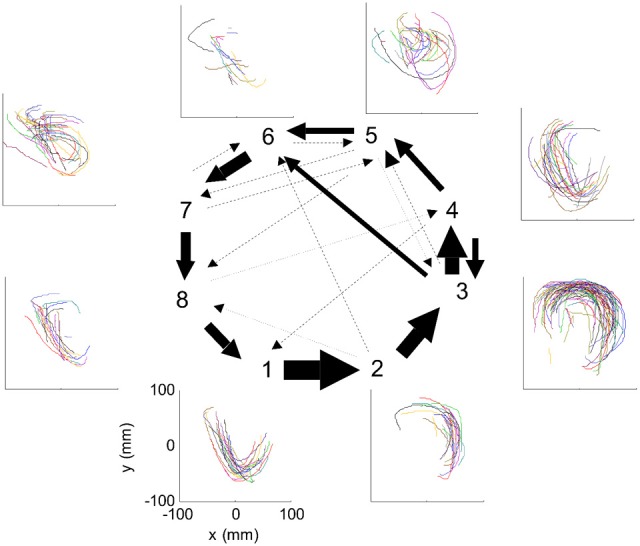
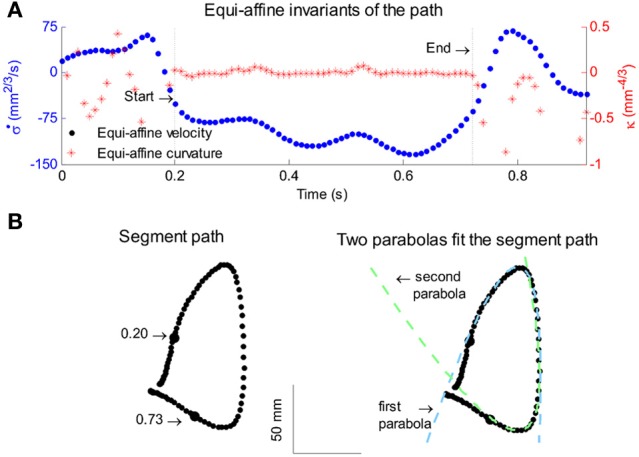
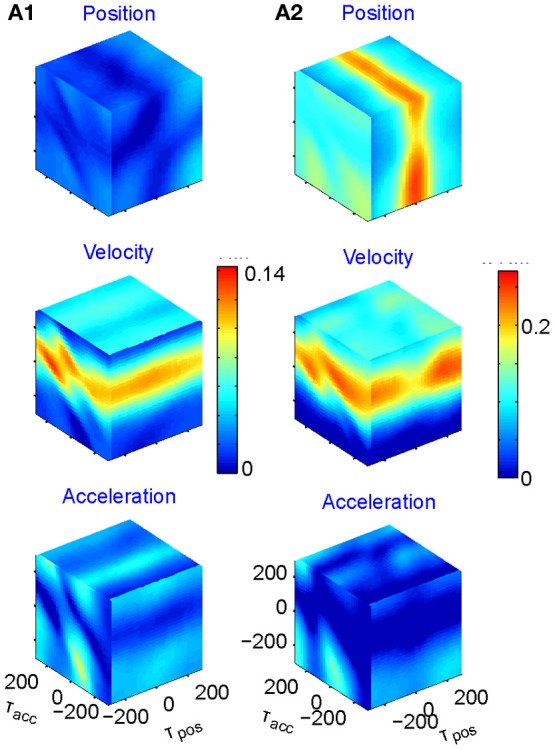
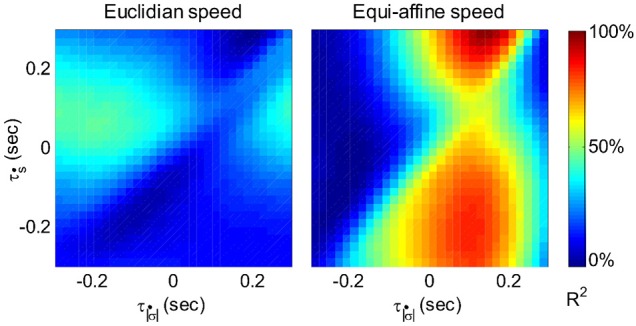
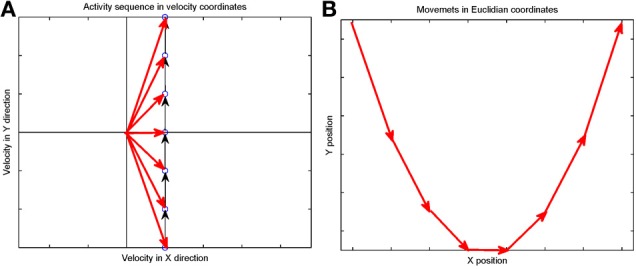
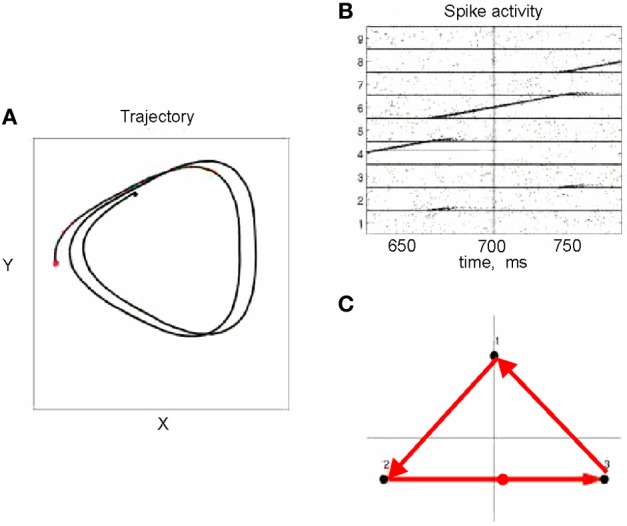
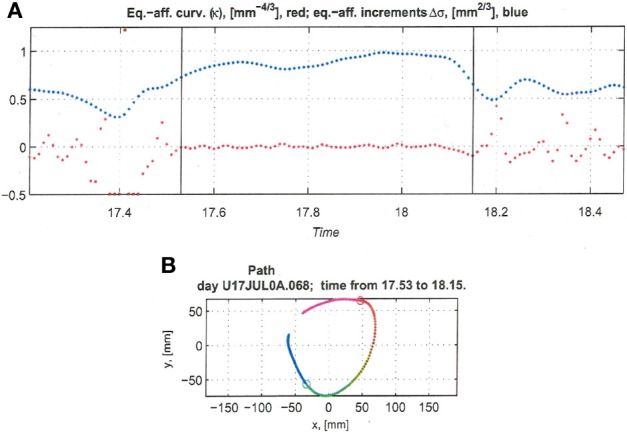
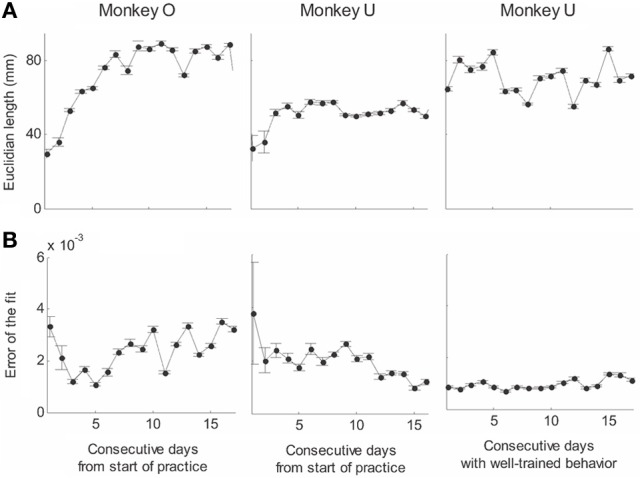
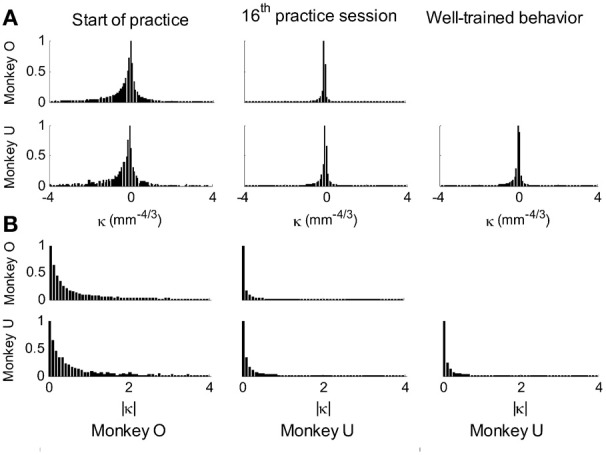
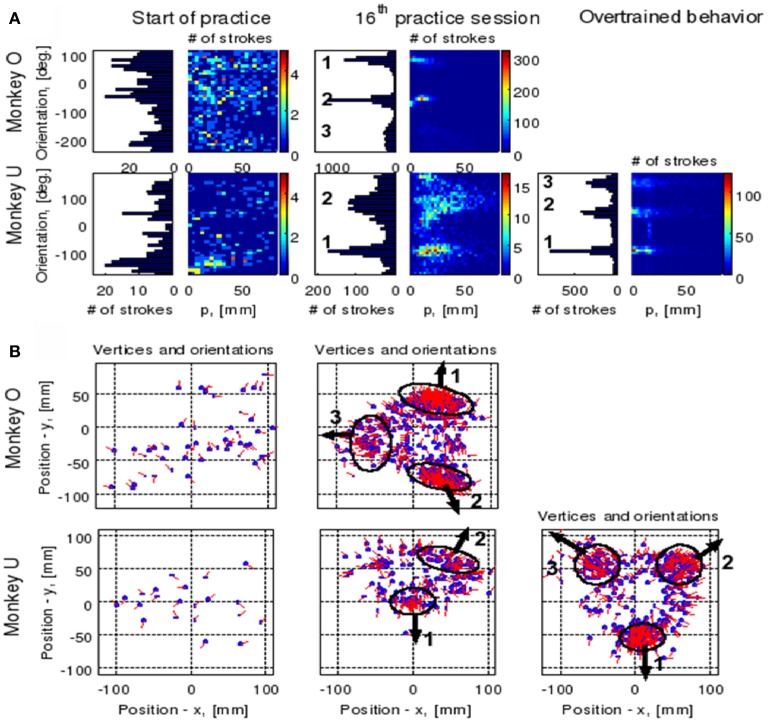
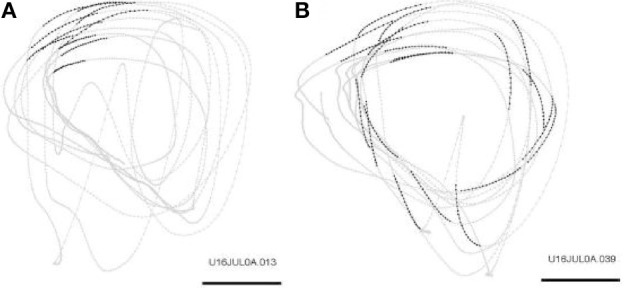
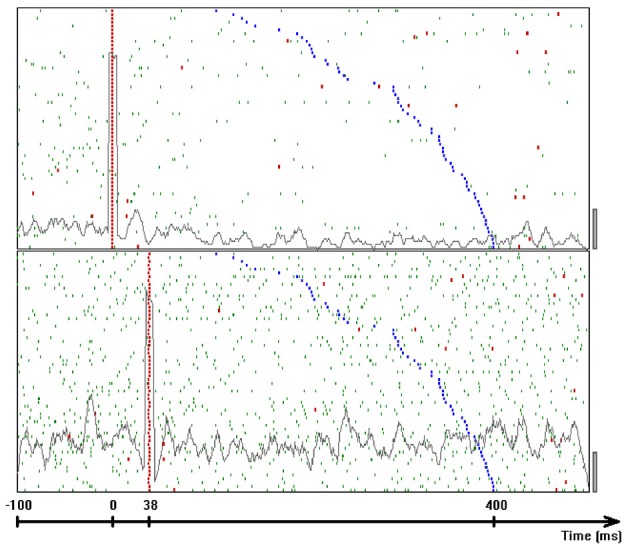
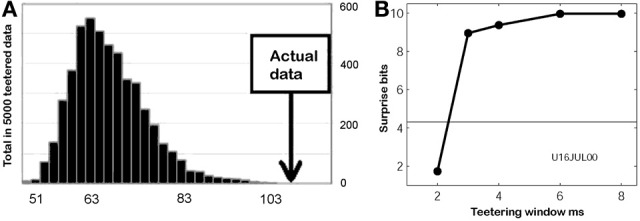
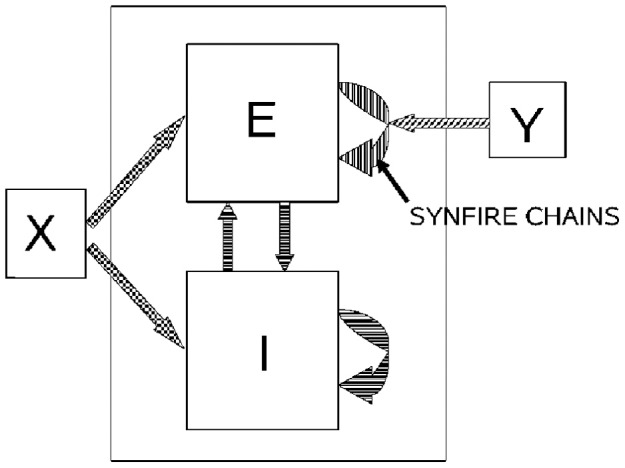
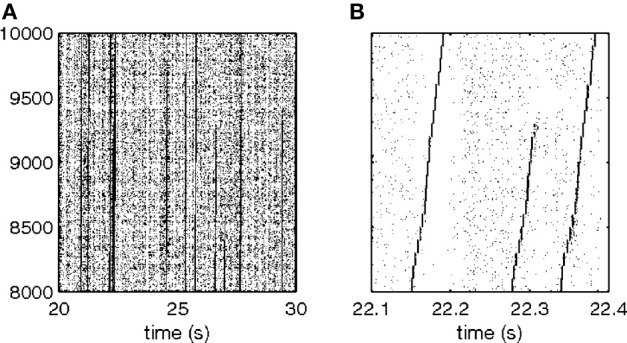
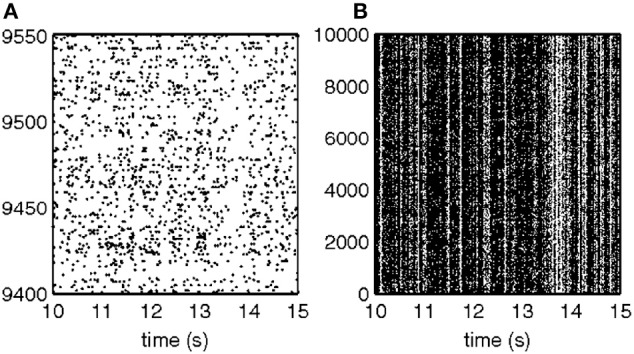
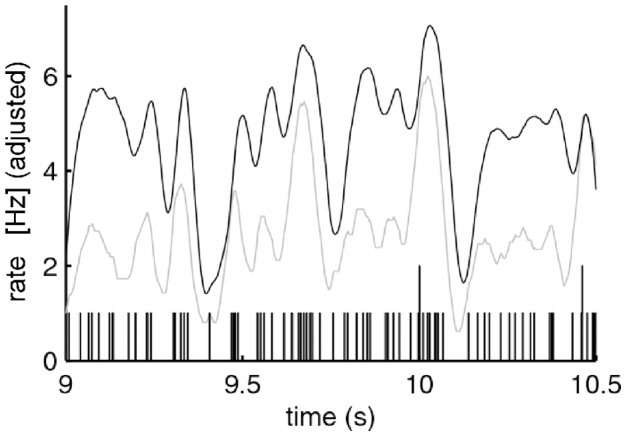
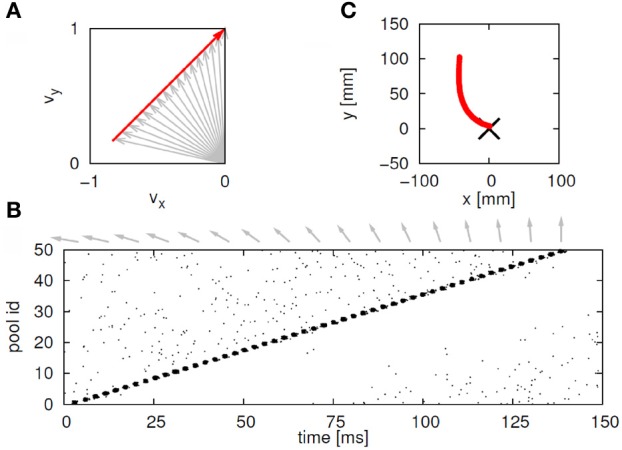
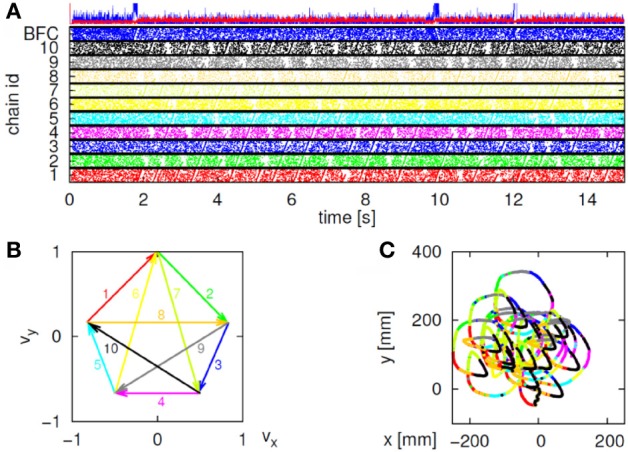
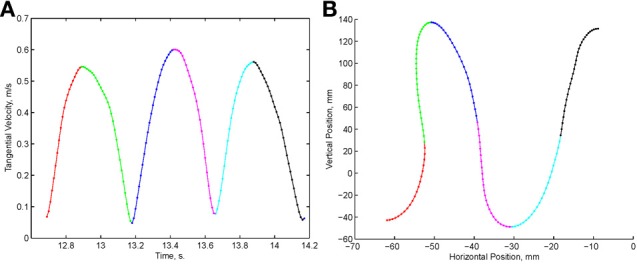
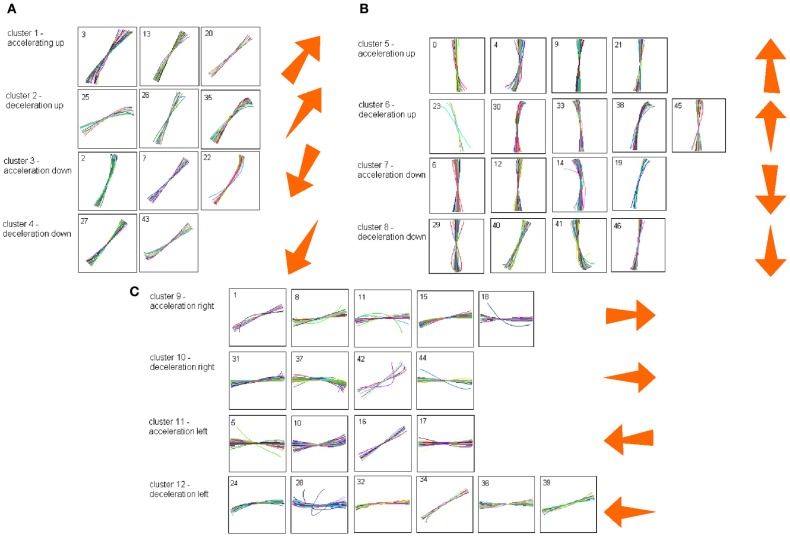
This article discusses the compositional structure of hand movements by analyzing and modeling neural and behavioral data obtained from experiments where a monkey (Macaca fascicularis) performed scribbling movements induced by a search task. Using geometrically based approaches to movement segmentation, it is shown that the hand trajectories are composed of elementary segments that are primarily parabolic in shape. The segments could be categorized into a small number of classes on the basis of decreasing intra-class variance over the course of training. A separate classification of the neural data employing a hidden Markov model showed a coincidence of the neural states with the behavioral categories. An additional analysis of both types of data by a data mining method provided evidence that the neural activity patterns underlying the behavioral primitives were formed by sets of specific and precise spike patterns. A geometric description of the movement trajectories, together with precise neural timing data indicates a compositional variant of a realistic synfire chain model. This model reproduces the typical shapes and temporal properties of the trajectories; hence the structure and composition of the primitives may reflect meaningful behavior.
Keywords: compositionality; hand-motion-model; motion-primitives; scribbling; synfire chains; voluntary-movements.
Figures
Similar articles
-
Compositionality of arm movements can be realized by propagating synchrony.J Comput Neurosci. 2011 Jun;30(3):675-97. doi: 10.1007/s10827-010-0285-9. Epub 2010 Oct 16. J Comput Neurosci. 2011. PMID: 20953686 Free PMC article.
-
A compact representation of drawing movements with sequences of parabolic primitives.PLoS Comput Biol. 2009 Jul;5(7):e1000427. doi: 10.1371/journal.pcbi.1000427. Epub 2009 Jul 3. PLoS Comput Biol. 2009. PMID: 19578429 Free PMC article.
-
A compositionality machine realized by a hierarchic architecture of synfire chains.Front Comput Neurosci. 2011 Jan 5;4:154. doi: 10.3389/fncom.2010.00154. eCollection 2011. Front Comput Neurosci. 2011. PMID: 21258641 Free PMC article.
-
Motor primitives in vertebrates and invertebrates.Curr Opin Neurobiol. 2005 Dec;15(6):660-6. doi: 10.1016/j.conb.2005.10.011. Epub 2005 Nov 4. Curr Opin Neurobiol. 2005. PMID: 16275056 Review.
-
Roles of primate spinal interneurons in preparation and execution of voluntary hand movement.Brain Res Brain Res Rev. 2002 Oct;40(1-3):53-65. doi: 10.1016/s0165-0173(02)00188-1. Brain Res Brain Res Rev. 2002. PMID: 12589906 Review.
Cited by
-
Analysis of complex neural circuits with nonlinear multidimensional hidden state models.Proc Natl Acad Sci U S A. 2016 Jun 7;113(23):6538-43. doi: 10.1073/pnas.1606280113. Epub 2016 May 24. Proc Natl Acad Sci U S A. 2016. PMID: 27222584 Free PMC article.
-
The speed-curvature power law of movements: a reappraisal.Exp Brain Res. 2018 Jan;236(1):69-82. doi: 10.1007/s00221-017-5108-z. Epub 2017 Oct 25. Exp Brain Res. 2018. PMID: 29071361
-
Quantifying spontaneous infant movements using state-space models.Sci Rep. 2024 Nov 19;14(1):28598. doi: 10.1038/s41598-024-80202-x. Sci Rep. 2024. PMID: 39562837 Free PMC article.
-
Biological kinematics: a detailed review of the velocity-curvature power law calculation.Exp Brain Res. 2025 Apr 3;243(5):107. doi: 10.1007/s00221-025-07065-0. Exp Brain Res. 2025. PMID: 40178611 Free PMC article. Review.
-
Motor primitives--new data and future questions.Curr Opin Neurobiol. 2015 Aug;33:156-65. doi: 10.1016/j.conb.2015.04.004. Epub 2015 Apr 22. Curr Opin Neurobiol. 2015. PMID: 25912883 Free PMC article. Review.
References
-
- Abeles M. (1982). Local Cortical Circuits: an Electrophysiological Study. Berlin; New York: Springer-Verlag
-
- Abeles M. (1991). Corticonics: Neural Circuits of the Cerebral Cortex. New York, NY: Cambridge University Press; 10.1017/CBO9780511574566 - DOI
-
- Abeles M., Goldstein M. H. (1977). Multispike train analysis. Proc. IEEE 65, 762–773 10.1109/PROC.1977.10559 - DOI
-
- Amarasingham A., Hastopoulos N., Geman S. (2003). At what time scale does the nervous system operate? Neurocomputing 52, 25–29
LinkOut - more resources
Full Text Sources
Other Literature Sources