

NDV-3 protects mice from vulvovaginal candidiasis through T- and B-cell immune response
- PMID: 24063977
- PMCID: PMC3866209
- DOI: 10.1016/j.vaccine.2013.09.016
NDV-3 protects mice from vulvovaginal candidiasis through T- and B-cell immune response
Abstract
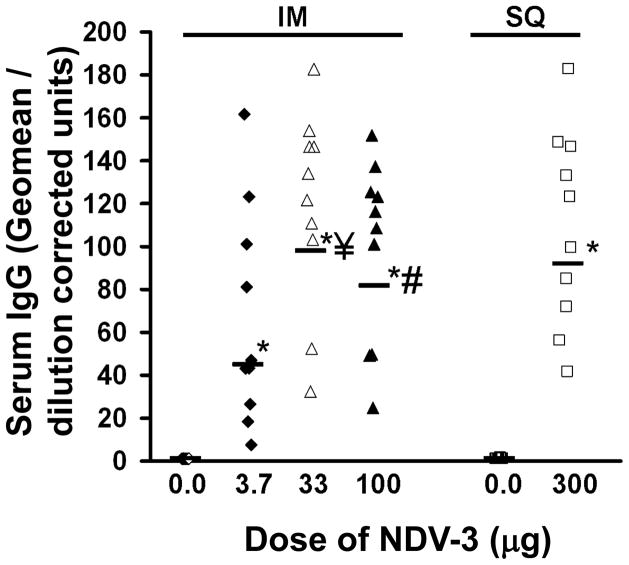
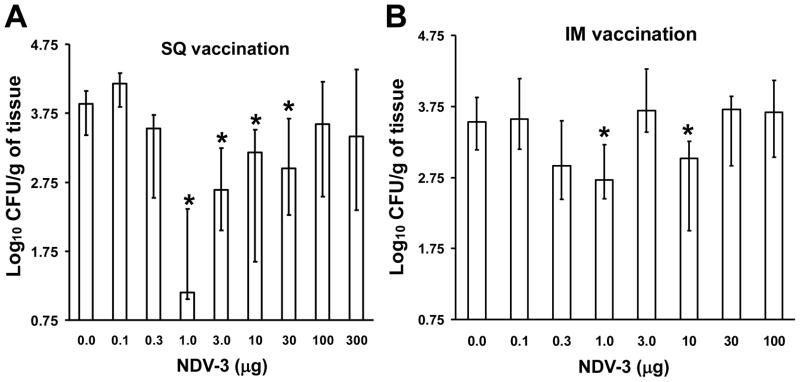
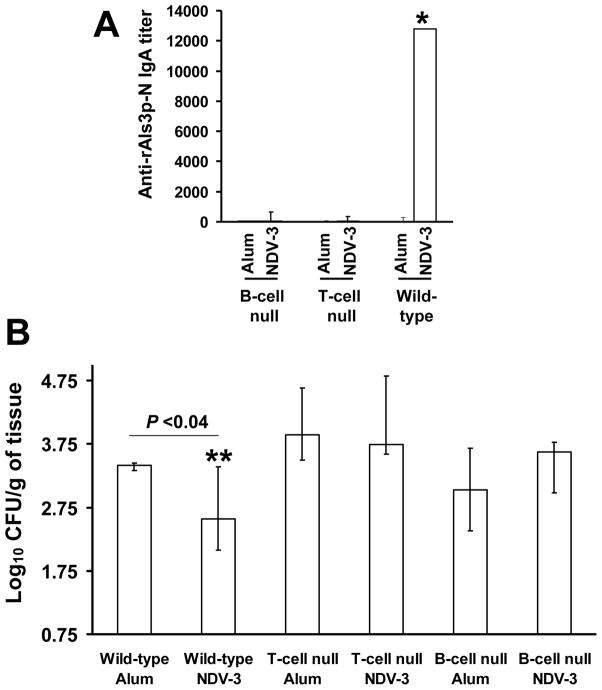
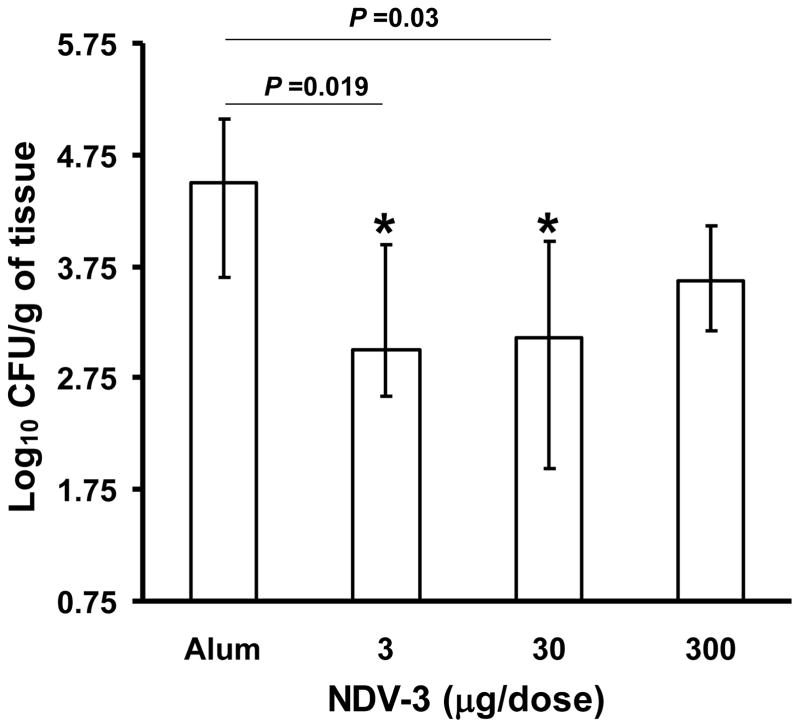
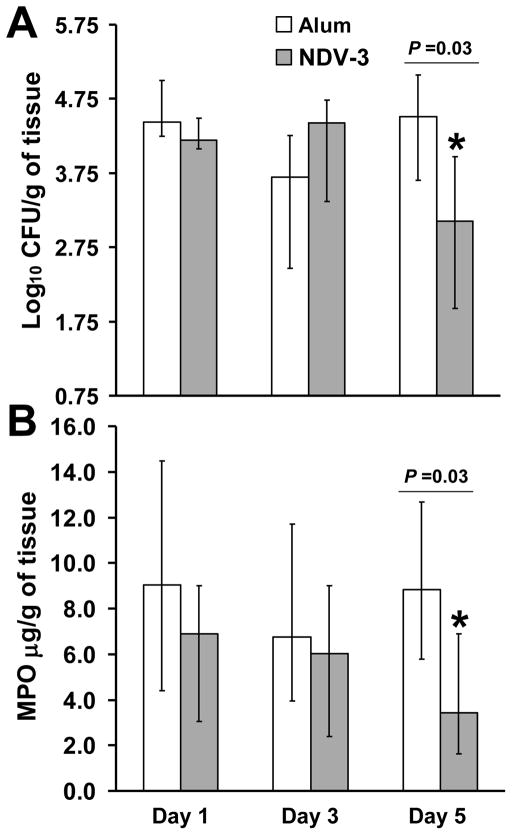
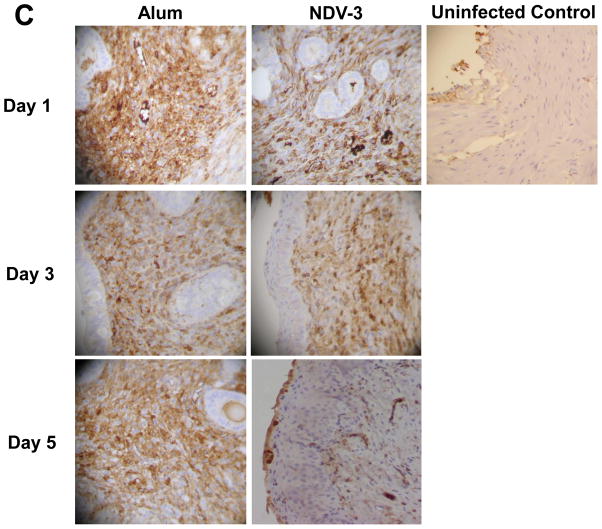
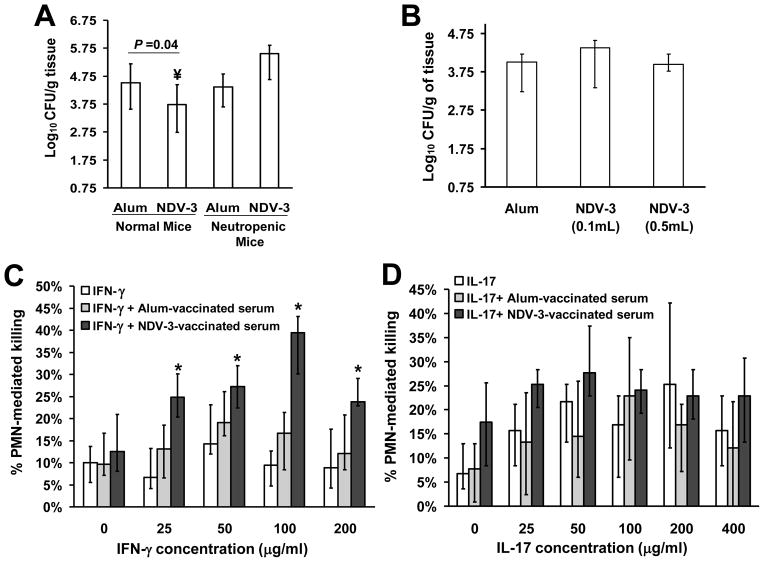
We have previously reported that vaccination with rAls3p-N protein of Candida albicans, formulated with alum adjuvant (also designated as NDV-3) protects immunocompetent mice from, lethal disseminated candidiasis and mucosal oropharyngeal candidiasis. NDV-3 vaccine was recently, tested in a Phase 1 clinical trial and found to be safe, well-tolerated, and induced robust humoral and, cellular immune responses with increased interferon (IFN)-gamma and interleukin (IL)-17 secretion. In preparation for a Phase 2 clinical trial against vulvovaginal candidiasis (VVC), we evaluated NDV-3, efficacy in a murine VVC model. Here, NDV-3 induced a strong immune response characterized by high, anti-rAls3p-N serum IgG and vaginal IgA titers. Furthermore, moderate doses of the vaccine (a range of 1-30μg given subcutaneously [SQ] or 0.3-10μg given intramuscularly [IM]) elicited a 10-1000 fold, decrease in vaginal fungal burden vs. control (mice injected with alum adjuvant alone) in both inbred, and outbred mice infected with different clinical C. albicans isolates. Additionally, NDV-3 required both, T and B lymphocytes for efficacy in reducing C. albicans tissue burden, which is followed by a reduction, in neutrophil influx to the affected site. Finally, anti-rAls3p-N antibodies enhanced the ex vivo killing, of C. albicans by neutrophils primed with IFN-gamma. These data indicate that NDV-3 protects mice, from VVC by a mechanism that involves the concerted priming of both humoral and adaptive immune, responses.
Keywords: Als3; Candida albicans; Murine; NDV-3; VVC.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
-
- Sobel JD, Hasegawa A, Debernardis F, Adriani D, Pellegrini G, Cassone A, et al. Selected animal models: vaginal candidosis, Pneumocystis pneumonia, dermatophytosis and trichosporosis. Med Mycol. 1998;36( Suppl 1):129–36. - PubMed
-
- Fidel PL, Jr, Sobel JD. Immunopathogenesis of recurrent vulvovaginal candidiasis. Contracept Fertil Sex. 1996 Jan;24(1):33–40. - PubMed
-
- Sobel JD, Wiesenfeld HC, Martens M, Danna P, Hooton TM, Rompalo A, et al. Maintenance fluconazole therapy for recurrent vulvovaginal candidiasis. N Engl J Med. 2004 Aug 26;351(9):876–83. - PubMed
-
- Bauters TG, Dhont MA, Temmerman MI, Nelis HJ. Prevalence of vulvovaginal candidiasis and susceptibility to fluconazole in women. Am J Obstet Gynecol. 2002 Sep;187(3):569–74. - PubMed
-
- Cernicka J, Subik J. Resistance mechanisms in fluconazole-resistant Candida albicans isolates from vaginal candidiasis. Int J Antimicrob Agents. 2006 May;27(5):403–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
