

Initiation of mRNA decay in bacteria
- PMID: 24064983
- PMCID: PMC3997798
- DOI: 10.1007/s00018-013-1472-4
Initiation of mRNA decay in bacteria
Abstract
The instability of messenger RNA is fundamental to the control of gene expression. In bacteria, mRNA degradation generally follows an "all-or-none" pattern. This implies that if control is to be efficient, it must occur at the initiating (and presumably rate-limiting) step of the degradation process. Studies of E. coli and B. subtilis, species separated by 3 billion years of evolution, have revealed the principal and very disparate enzymes involved in this process in the two organisms. The early view that mRNA decay in these two model organisms is radically different has given way to new models that can be resumed by "different enzymes-similar strategies". The recent characterization of key ribonucleases sheds light on an impressive case of convergent evolution that illustrates that the surprisingly similar functions of these totally unrelated enzymes are of general importance to RNA metabolism in bacteria. We now know that the major mRNA decay pathways initiate with an endonucleolytic cleavage in E. coli and B. subtilis and probably in many of the currently known bacteria for which these organisms are considered representative. We will discuss here the different pathways of eubacterial mRNA decay, describe the major players and summarize the events that can precede and/or favor nucleolytic inactivation of a mRNA, notably the role of the 5' end and translation initiation. Finally, we will discuss the role of subcellular compartmentalization of transcription, translation, and the RNA degradation machinery.
Figures
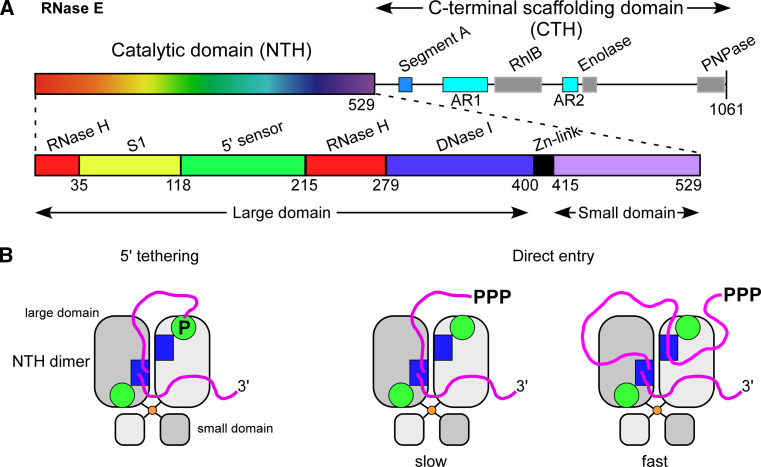
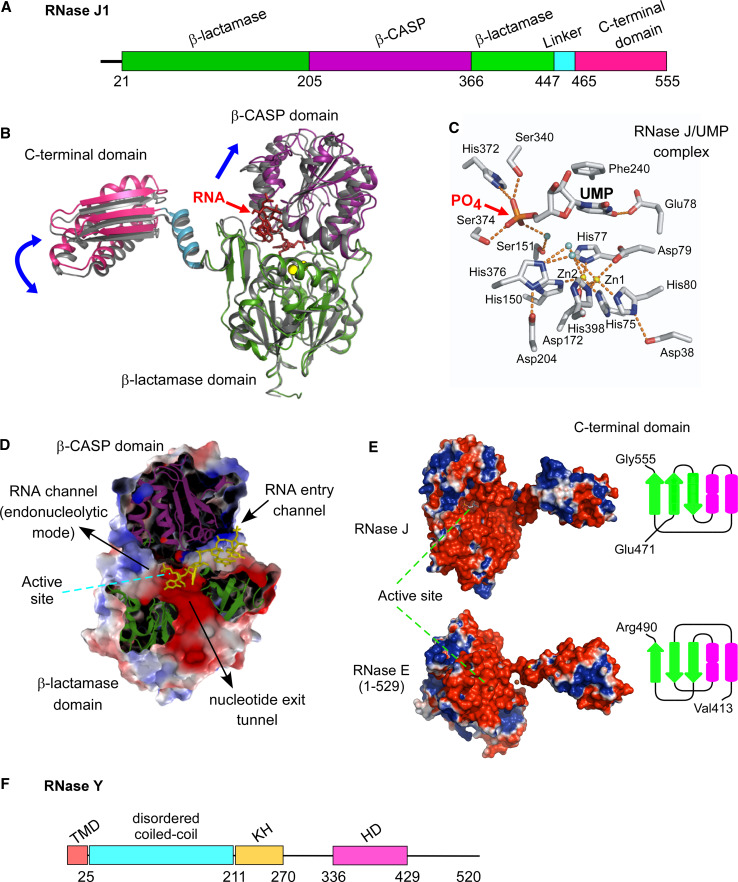
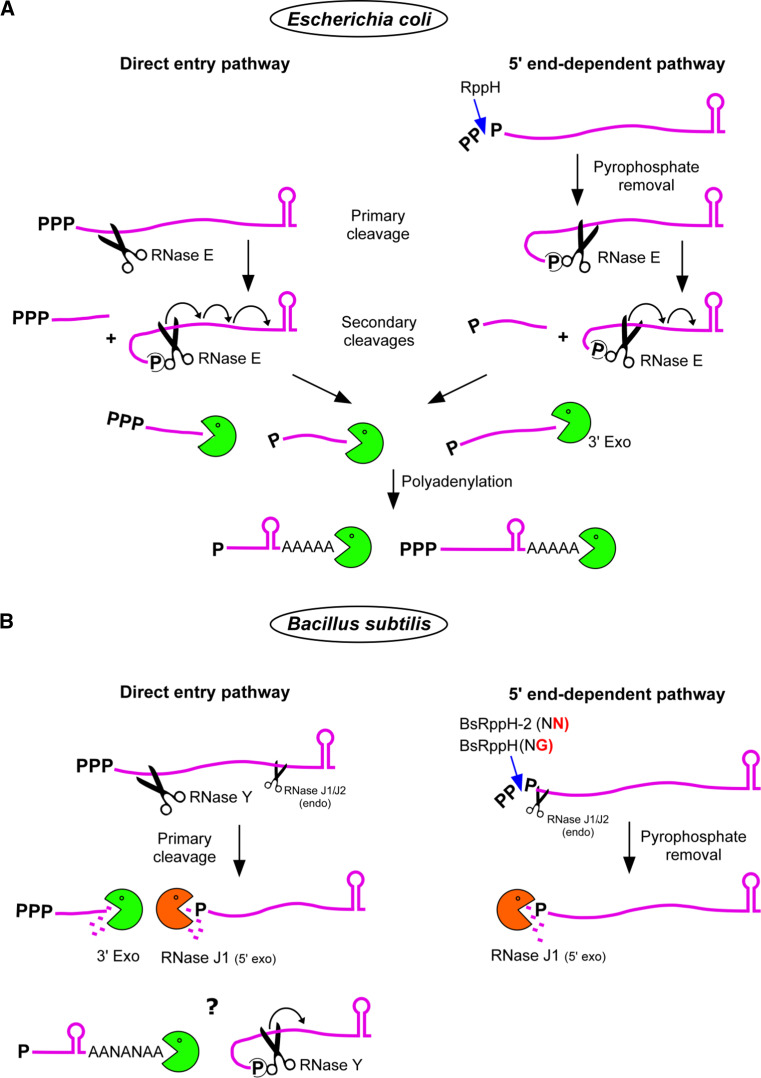
References
-
- Ludwig H, Homuth G, Schmalisch M, Dyka FM, Hecker M, Stulke J. Transcription of glycolytic genes and operons in Bacillus subtilis: evidence for the presence of multiple levels of control of the gapA operon. Mol Microbiol. 2001;41:409–422. - PubMed
-
- Marincola G, Schafer T, Behler J, Bernhardt J, Ohlsen K, Goerke C, Wolz C. RNase Y of Staphylococcus aureus and its role in the activation of virulence genes. Mol Microbiol. 2012;85:817–832. - PubMed
-
- Dreyfus M. Killer and protective ribosomes. Prog Mol Biol Transl Sci. 2009;85:423–466. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
