

H3.3 actively marks enhancers and primes gene transcription via opening higher-ordered chromatin
- PMID: 24065740
- PMCID: PMC3850095
- DOI: 10.1101/gad.222174.113
H3.3 actively marks enhancers and primes gene transcription via opening higher-ordered chromatin
Erratum in
-
Corrigendum: H3.3 actively marks enhancers and primes gene transcription via opening higher-ordered chromatin.Genes Dev. 2021 May 1;35(9-10):782. doi: 10.1101/gad.348504.121. Genes Dev. 2021. PMID: 33941541 Free PMC article. No abstract available.
Abstract
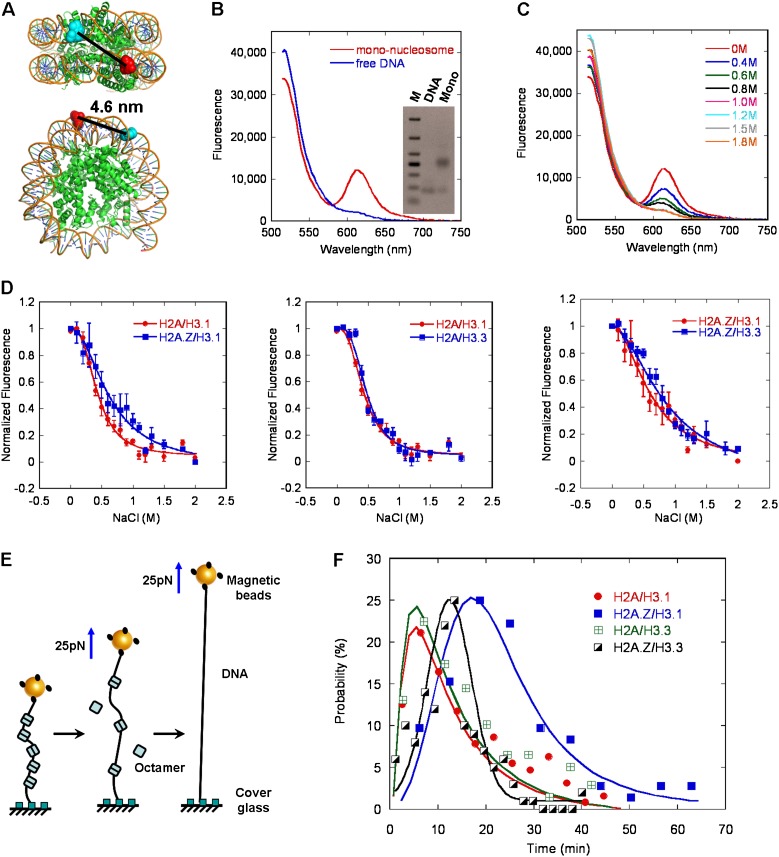
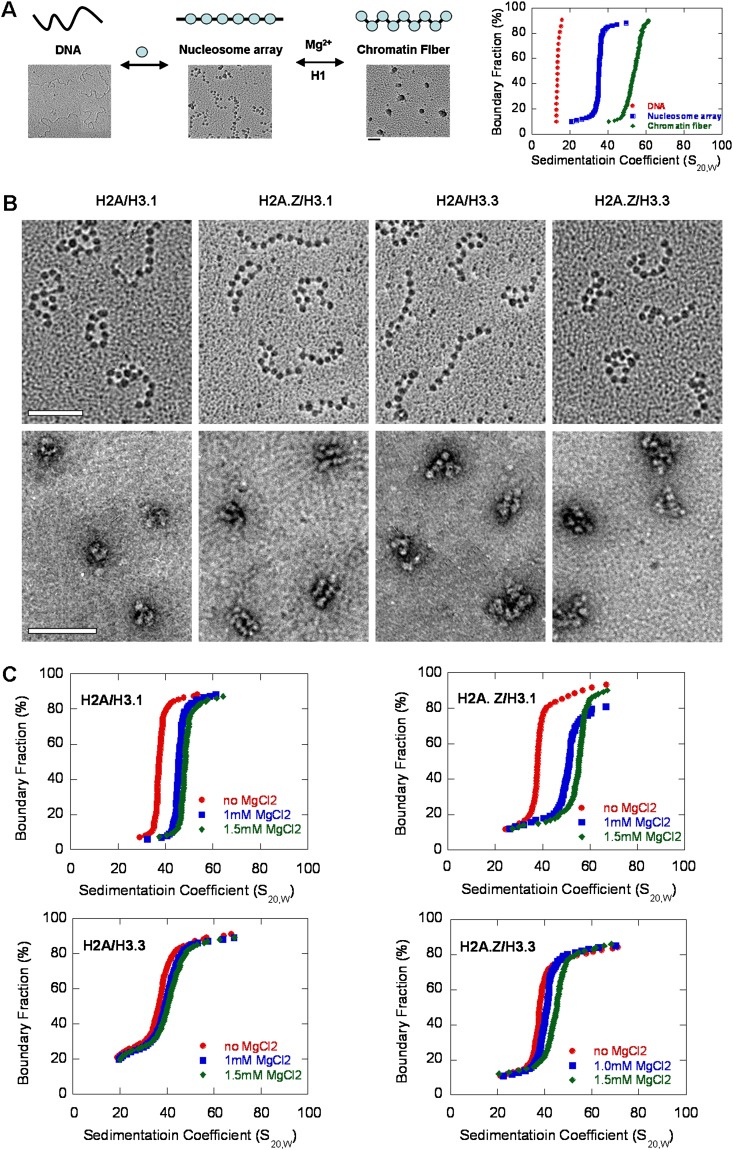
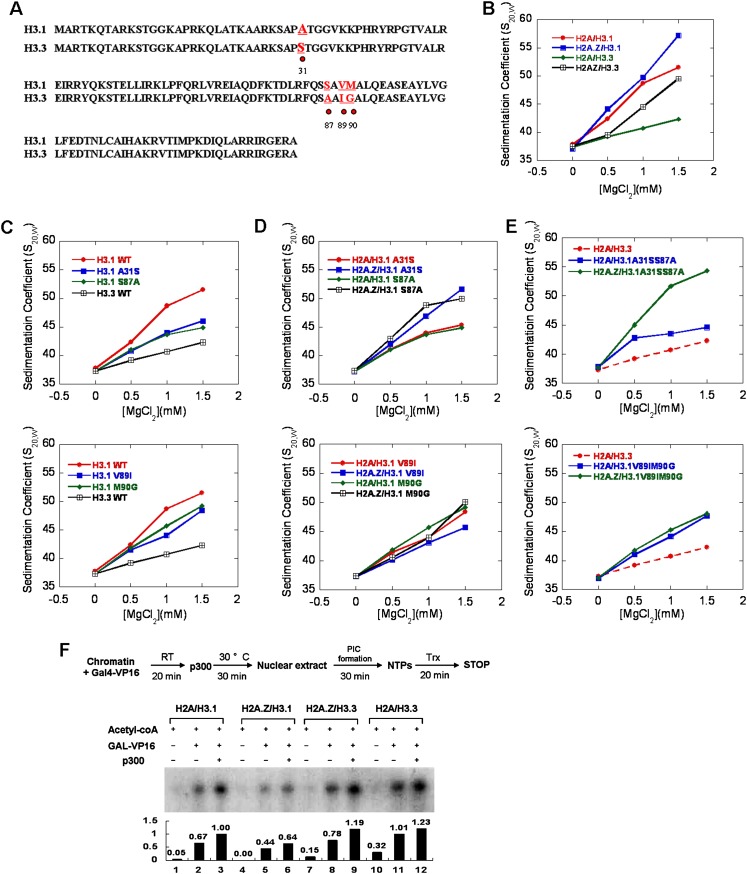
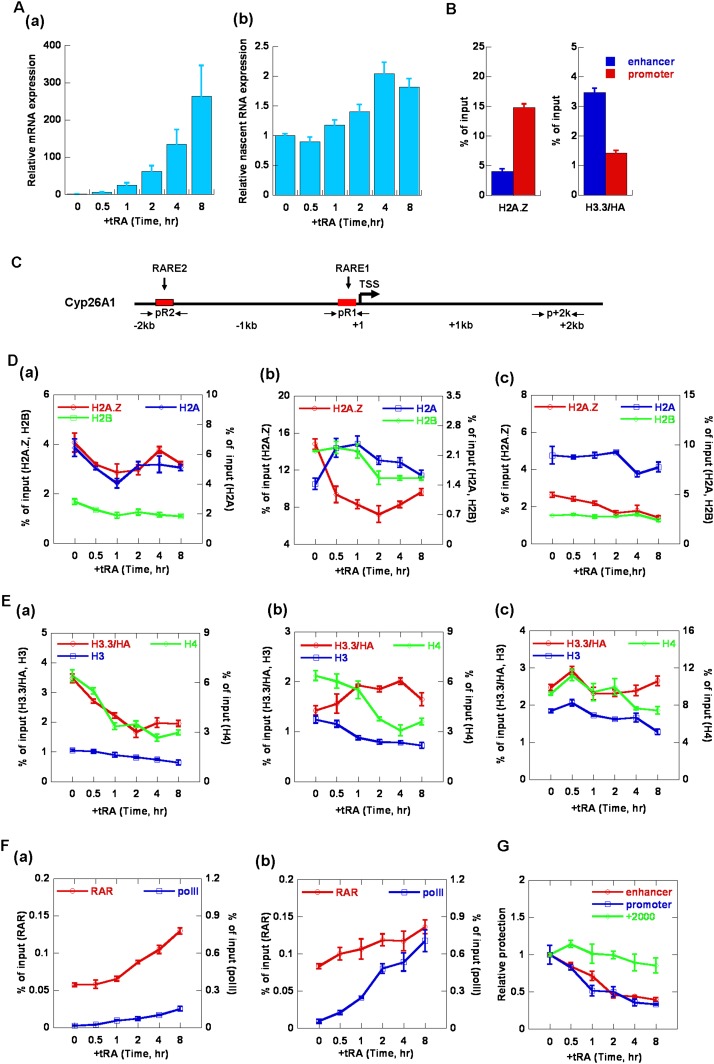
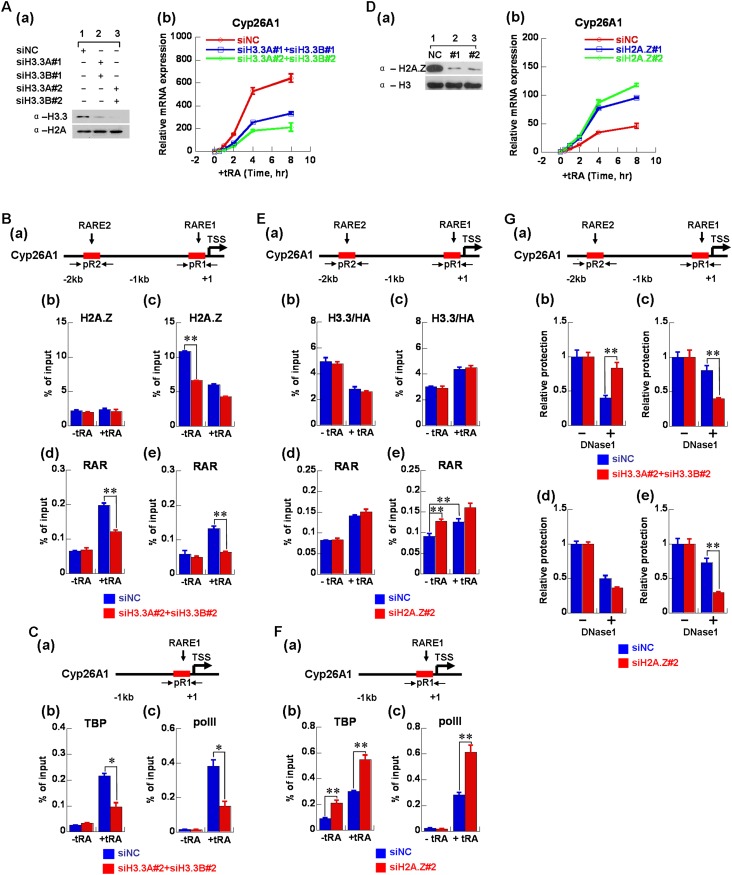
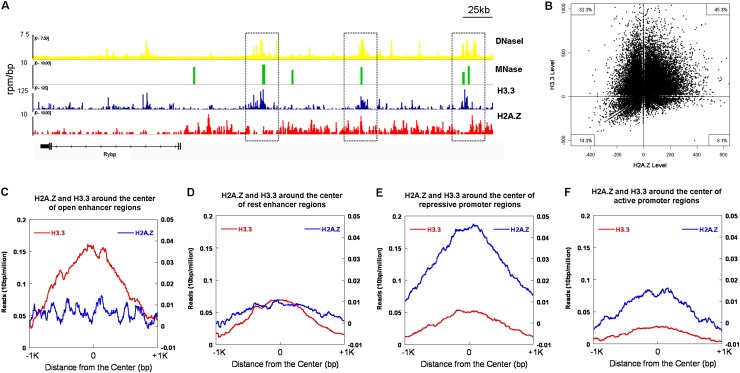
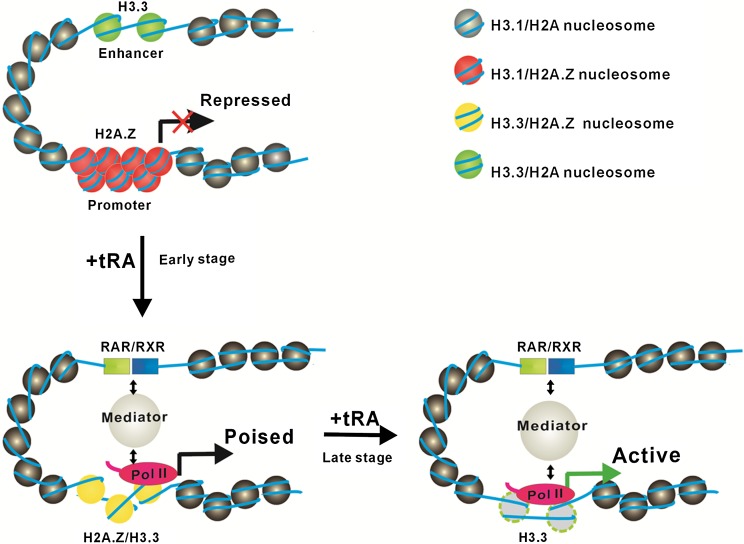
The histone variants H3.3 and H2A.Z have recently emerged as two of the most important features in transcriptional regulation, the molecular mechanism of which still remains poorly understood. In this study, we investigated the regulation of H3.3 and H2A.Z on chromatin dynamics during transcriptional activation. Our in vitro biophysical and biochemical investigation showed that H2A.Z promoted chromatin compaction and repressed transcriptional activity. Surprisingly, with only four to five amino acid differences from the canonical H3, H3.3 greatly impaired higher-ordered chromatin folding and promoted gene activation, although it has no significant effect on the stability of mononucleosomes. We further demonstrated that H3.3 actively marks enhancers and determines the transcriptional potential of retinoid acid (RA)-regulated genes via creating an open chromatin signature that enables the binding of RAR/RXR. Additionally, the H3.3-dependent recruitment of H2A.Z on promoter regions resulted in compaction of chromatin to poise transcription, while RA induction results in the incorporation of H3.3 on promoter regions to activate transcription via counteracting H2A.Z-mediated chromatin compaction. Our results provide key insights into the mechanism of how histone variants H3.3 and H2A.Z function together to regulate gene transcription via the modulation of chromatin dynamics over the enhancer and promoter regions.
Keywords: H2A.Z; H3.3; chromatin dynamics; gene transcription; higher-ordered chromatin structure; histone variants.
Figures
References
-
- Ahmad K, Henikoff S 2002. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol Cell 9: 1191–1200 - PubMed
-
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K 2007. High-resolution profiling of histone methylations in the human genome. Cell 129: 823–837 - PubMed
-
- Dalvai M, Bellucci L, Fleury L, Lavigne AC, Moutahir F, Bystricky K 2012. H2A.Z-dependent crosstalk between enhancer and promoter regulates Cyclin D1 expression. Oncogene 32: 4243–4251 - PubMed
-
- Dyer PN, Edayathumangalam RS, White CL, Bao Y, Chakravarthy S, Muthurajan UM, Luger K 2004. Reconstitution of nucleosome core particles from recombinant histones and DNA. Methods Enzymol 375: 23–44 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases