

Exchange of associated factors directs a switch in HBO1 acetyltransferase histone tail specificity
- PMID: 24065767
- PMCID: PMC3792477
- DOI: 10.1101/gad.223396.113
Exchange of associated factors directs a switch in HBO1 acetyltransferase histone tail specificity
Abstract
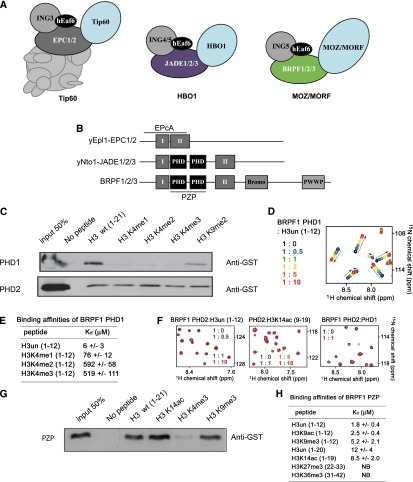
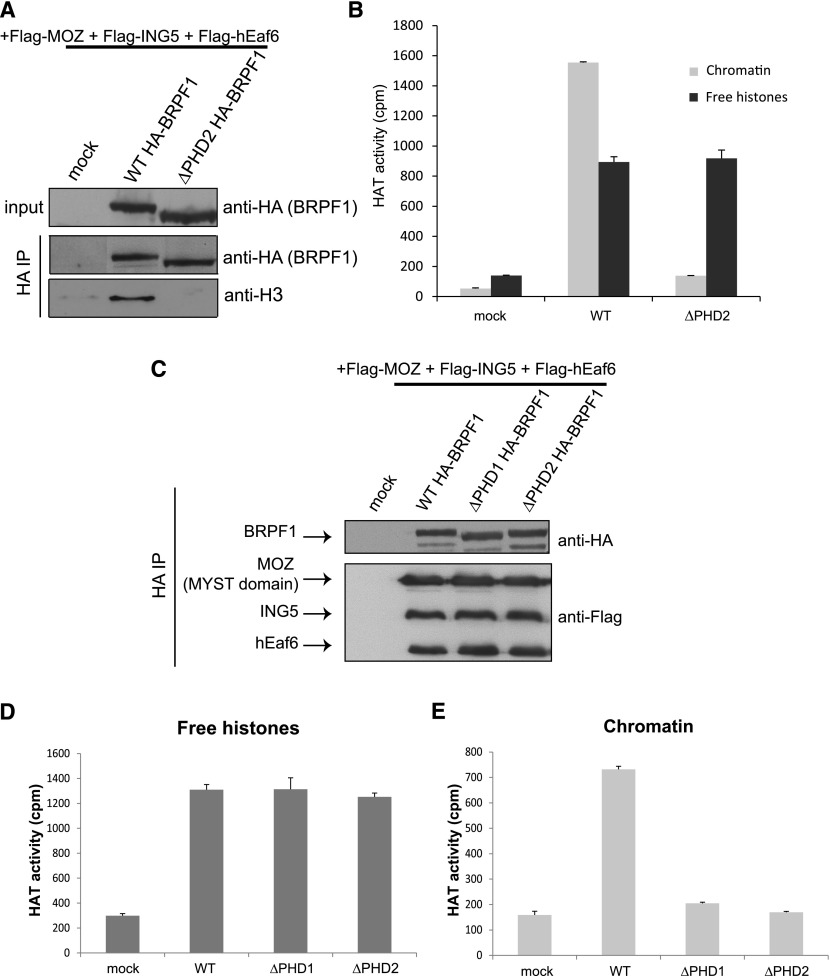
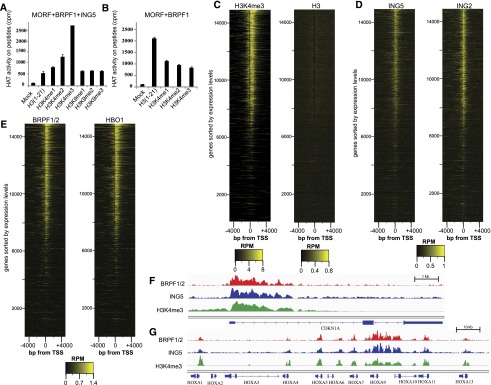
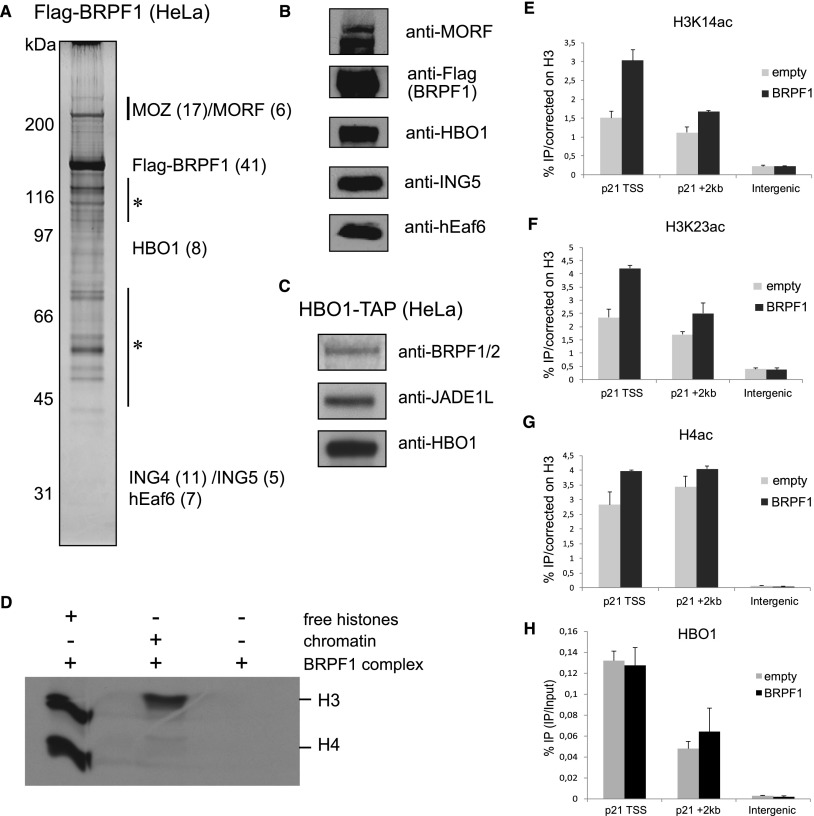
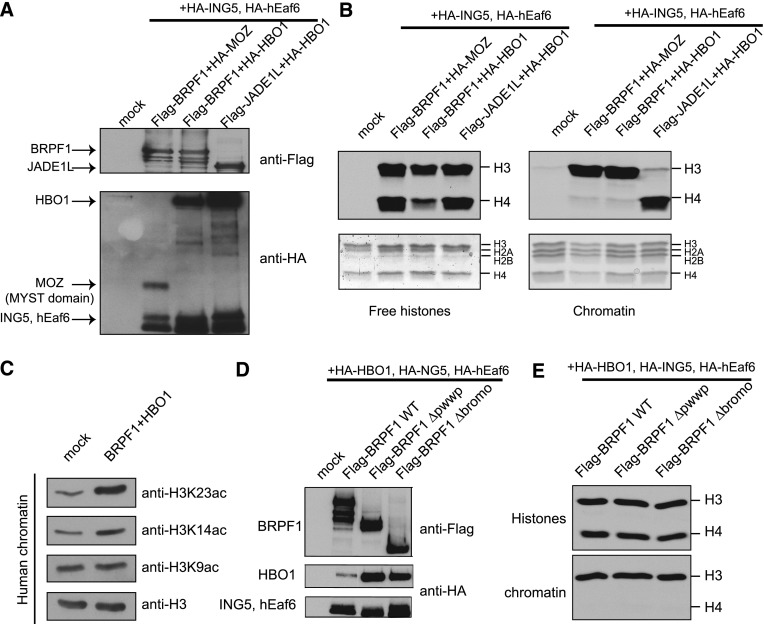
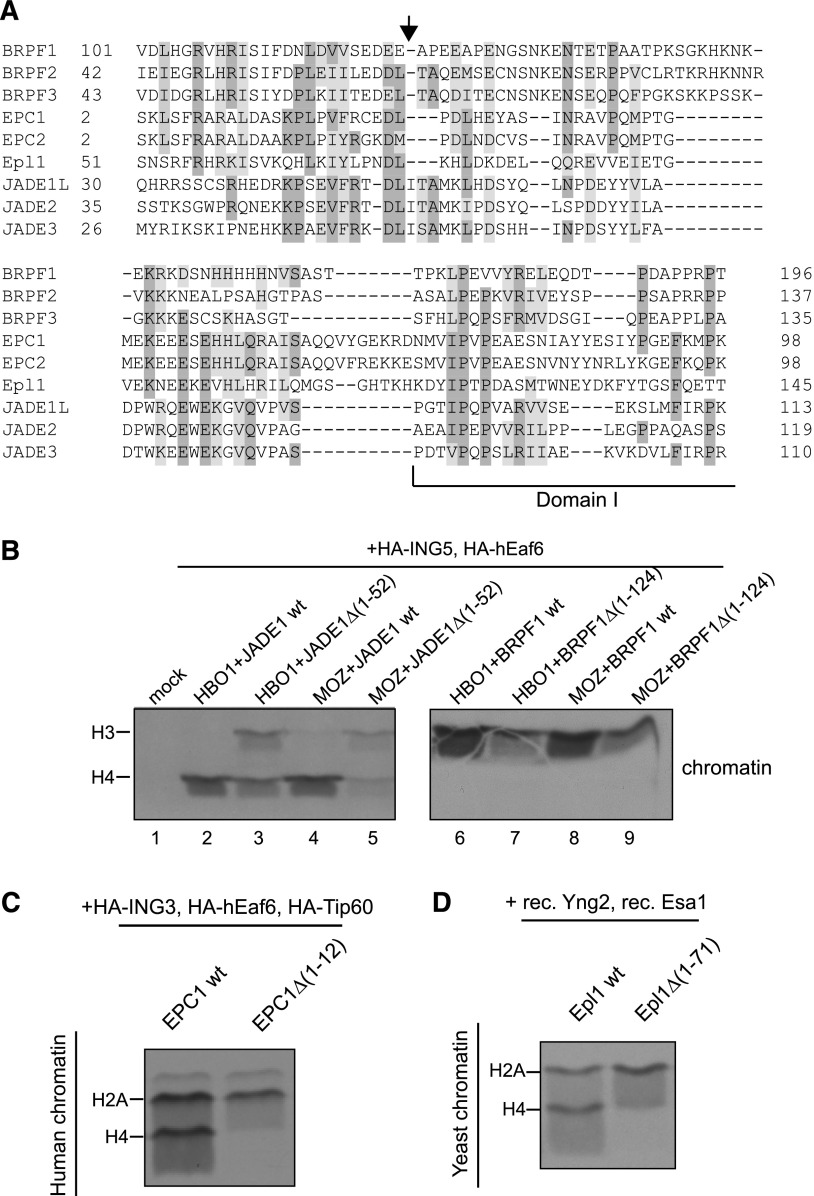
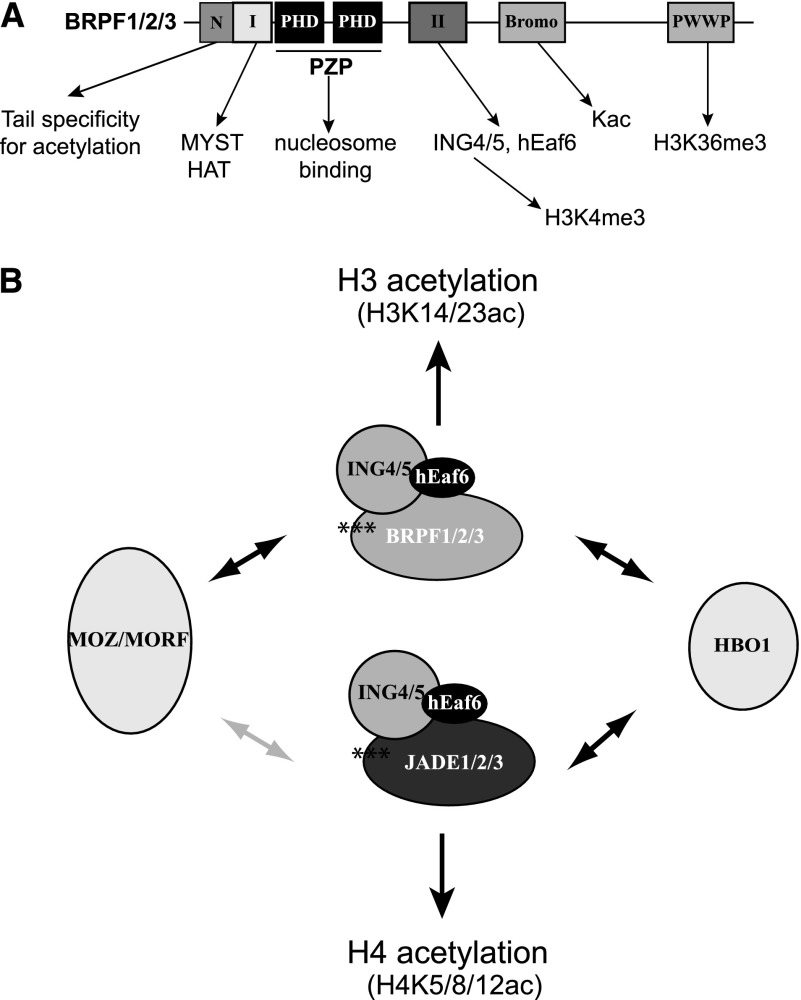
Histone acetyltransferases (HATs) assemble into multisubunit complexes in order to target distinct lysine residues on nucleosomal histones. Here, we characterize native HAT complexes assembled by the BRPF family of scaffold proteins. Their plant homeodomain (PHD)-Zn knuckle-PHD domain is essential for binding chromatin and is restricted to unmethylated H3K4, a specificity that is reversed by the associated ING subunit. Native BRPF1 complexes can contain either MOZ/MORF or HBO1 as catalytic acetyltransferase subunit. Interestingly, while the previously reported HBO1 complexes containing JADE scaffold proteins target histone H4, the HBO1-BRPF1 complex acetylates only H3 in chromatin. We mapped a small region to the N terminus of scaffold proteins responsible for histone tail selection on chromatin. Thus, alternate choice of subunits associated with HBO1 can switch its specificity between H4 and H3 tails. These results uncover a crucial new role for associated proteins within HAT complexes, previously thought to be intrinsic to the catalytic subunit.
Keywords: BRPF1; MYST family; PHD fingers; acetyltransferase complexes; chromatin acetylation; histone tails.
Figures
Similar articles
-
The scaffolding protein JADE1 physically links the acetyltransferase subunit HBO1 with its histone H3-H4 substrate.J Biol Chem. 2018 Mar 23;293(12):4498-4509. doi: 10.1074/jbc.RA117.000677. Epub 2018 Jan 30. J Biol Chem. 2018. PMID: 29382722 Free PMC article.
-
Structural and mechanistic insights into regulation of HBO1 histone acetyltransferase activity by BRPF2.Nucleic Acids Res. 2017 Jun 2;45(10):5707-5719. doi: 10.1093/nar/gkx142. Nucleic Acids Res. 2017. PMID: 28334966 Free PMC article.
-
ING tumor suppressor proteins are critical regulators of chromatin acetylation required for genome expression and perpetuation.Mol Cell. 2006 Jan 6;21(1):51-64. doi: 10.1016/j.molcel.2005.12.007. Mol Cell. 2006. PMID: 16387653
-
Deciphering structure, function and mechanism of lysine acetyltransferase HBO1 in protein acetylation, transcription regulation, DNA replication and its oncogenic properties in cancer.Cell Mol Life Sci. 2020 Feb;77(4):637-649. doi: 10.1007/s00018-019-03296-x. Epub 2019 Sep 18. Cell Mol Life Sci. 2020. PMID: 31535175 Free PMC article. Review.
-
HBO1, a MYSTerious KAT and its links to cancer.Biochim Biophys Acta Gene Regul Mech. 2024 Sep;1867(3):195045. doi: 10.1016/j.bbagrm.2024.195045. Epub 2024 Jun 6. Biochim Biophys Acta Gene Regul Mech. 2024. PMID: 38851533 Review.
Cited by
-
The chromatin regulator Brpf1 regulates embryo development and cell proliferation.J Biol Chem. 2015 May 1;290(18):11349-64. doi: 10.1074/jbc.M115.643189. Epub 2015 Mar 15. J Biol Chem. 2015. PMID: 25773539 Free PMC article.
-
Regulation of KAT6 Acetyltransferases and Their Roles in Cell Cycle Progression, Stem Cell Maintenance, and Human Disease.Mol Cell Biol. 2016 Jun 29;36(14):1900-7. doi: 10.1128/MCB.00055-16. Print 2016 Jul 15. Mol Cell Biol. 2016. PMID: 27185879 Free PMC article. Review.
-
BRPF1 is essential for development of fetal hematopoietic stem cells.J Clin Invest. 2016 Sep 1;126(9):3247-62. doi: 10.1172/JCI80711. Epub 2016 Aug 8. J Clin Invest. 2016. PMID: 27500495 Free PMC article.
-
Molecular Basis for the PZP Domain of BRPF1 Association with Chromatin.Structure. 2020 Jan 7;28(1):105-110.e3. doi: 10.1016/j.str.2019.10.014. Epub 2019 Nov 8. Structure. 2020. PMID: 31711755 Free PMC article.
-
UHRF1-KAT7-mediated regulation of TUSC3 expression via histone methylation/acetylation is critical for the proliferation of colon cancer cells.Oncogene. 2020 Jan;39(5):1018-1030. doi: 10.1038/s41388-019-1032-y. Epub 2019 Oct 3. Oncogene. 2020. PMID: 31582837
References
-
- Abmayr SM, Yao T, Parmely T, Workman JL 1993. Preparation of nuclear and cytoplasmic extracts from mammalian cells. Curr Protoc Mol Biol 12: 12.1.1–12.1.9 - PubMed
-
- Agalioti T, Chen G, Thanos D 2002. Deciphering the transcriptional histone acetylation code for a human gene. Cell 111: 381–392 - PubMed
-
- Agricola E, Verdone L, Di Mauro E, Caserta M 2006. H4 acetylation does not replace H3 acetylation in chromatin remodelling and transcription activation of Adr1-dependent genes. Mol Microbiol 62: 1433–1446 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases