

De- and repolarization mechanism of flagellar morphogenesis during a bacterial cell cycle
- PMID: 24065770
- PMCID: PMC3792480
- DOI: 10.1101/gad.222679.113
De- and repolarization mechanism of flagellar morphogenesis during a bacterial cell cycle
Erratum in
- Genes Dev. 2013 Oct 15;27(20):2292
Abstract
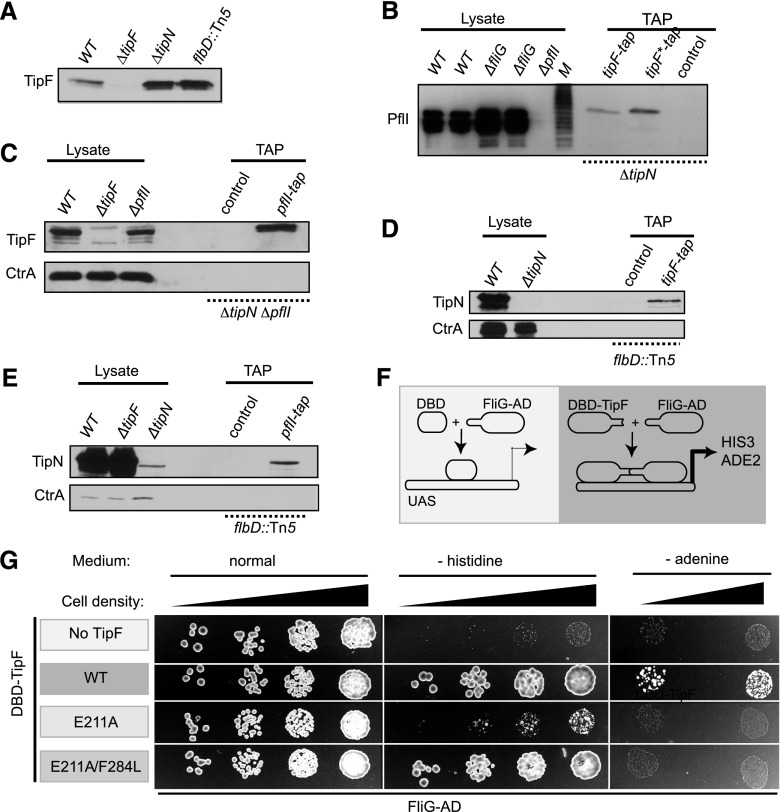
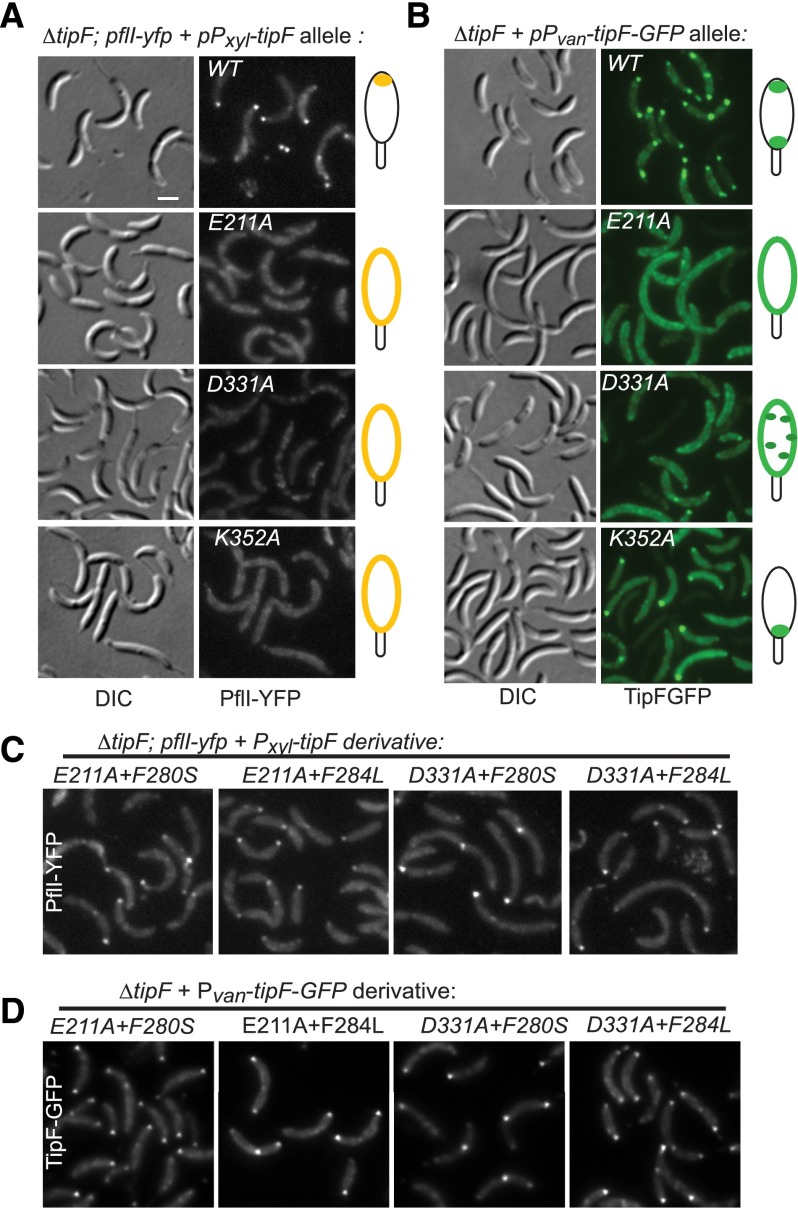
Eukaryotic morphogenesis is seeded with the establishment and subsequent amplification of polarity cues at key times during the cell cycle, often using (cyclic) nucleotide signals. We discovered that flagellum de- and repolarization in the model prokaryote Caulobacter crescentus is precisely orchestrated through at least three spatiotemporal mechanisms integrated at TipF. We show that TipF is a cell cycle-regulated receptor for the second messenger--bis-(3'-5')-cyclic dimeric guanosine monophosphate (c-di-GMP)--that perceives and transduces this signal through the degenerate c-di-GMP phosphodiesterase (EAL) domain to nucleate polar flagellum biogenesis. Once c-di-GMP levels rise at the G1 → S transition, TipF is activated, stabilized, and polarized, enabling the recruitment of downstream effectors, including flagellar switch proteins and the PflI positioning factor, at a preselected pole harboring the TipN landmark. These c-di-GMP-dependent events are coordinated with the onset of tipF transcription in early S phase and together enable the correct establishment and robust amplification of TipF-dependent polarization early in the cell cycle. Importantly, these mechanisms also govern the timely removal of TipF at cell division coincident with the drop in c-di-GMP levels, thereby resetting the flagellar polarization state in the next cell cycle after a preprogrammed period during which motility must be suspended.
Keywords: Caulobacter; asymmetric division; c-di-GMP; cell cycle; flagellum; morphogenesis; polarity.
Figures





Similar articles
-
Bacterial birth scar proteins mark future flagellum assembly site.Cell. 2006 Mar 10;124(5):1025-37. doi: 10.1016/j.cell.2006.01.019. Cell. 2006. PMID: 16530048
-
Probing flagellar promoter occupancy in wild-type and mutant Caulobacter crescentus by chromatin immunoprecipitation.FEMS Microbiol Lett. 2011 Jun;319(2):146-52. doi: 10.1111/j.1574-6968.2011.02275.x. Epub 2011 Apr 19. FEMS Microbiol Lett. 2011. PMID: 21457294
-
Precise timing of transcription by c-di-GMP coordinates cell cycle and morphogenesis in Caulobacter.Nat Commun. 2020 Feb 10;11(1):816. doi: 10.1038/s41467-020-14585-6. Nat Commun. 2020. PMID: 32041947 Free PMC article.
-
Interplay between flagellation and cell cycle control in Caulobacter.Curr Opin Microbiol. 2015 Dec;28:83-92. doi: 10.1016/j.mib.2015.08.012. Epub 2015 Oct 23. Curr Opin Microbiol. 2015. PMID: 26476805 Review.
-
Taming the flagellar motor of pseudomonads with a nucleotide messenger.Environ Microbiol. 2020 Jul;22(7):2496-2513. doi: 10.1111/1462-2920.15036. Epub 2020 May 5. Environ Microbiol. 2020. PMID: 32329141 Review.
Cited by
-
Proteolysis dependent cell cycle regulation in Caulobacter crescentus.Cell Div. 2022 Apr 1;17(1):3. doi: 10.1186/s13008-022-00078-z. Cell Div. 2022. PMID: 35365160 Free PMC article. Review.
-
Functional diversity of c-di-GMP receptors in prokaryotic and eukaryotic systems.Cell Commun Signal. 2023 Sep 25;21(1):259. doi: 10.1186/s12964-023-01263-5. Cell Commun Signal. 2023. PMID: 37749602 Free PMC article. Review.
-
Polar confinement of a macromolecular machine by an SRP-type GTPase.Nat Commun. 2024 Jul 10;15(1):5797. doi: 10.1038/s41467-024-50274-4. Nat Commun. 2024. PMID: 38987236 Free PMC article.
-
Contribution of Physical Interactions to Signaling Specificity between a Diguanylate Cyclase and Its Effector.mBio. 2015 Dec 15;6(6):e01978-15. doi: 10.1128/mBio.01978-15. mBio. 2015. PMID: 26670387 Free PMC article.
-
We're in this Together: Sensation of the Host Cell Environment by Endosymbiotic Bacteria.Results Probl Cell Differ. 2020;69:179-197. doi: 10.1007/978-3-030-51849-3_6. Results Probl Cell Differ. 2020. PMID: 33263872
References
-
- Alley MR, Maddock JR, Shapiro L 1992. Polar localization of a bacterial chemoreceptor. Genes Dev 6: 825–836 - PubMed
-
- Barends TR, Hartmann E, Griese JJ, Beitlich T, Kirienko NV, Ryjenkov DA, Reinstein J, Shoeman RL, Gomelsky M, Schlichting I 2009. Structure and mechanism of a bacterial light-regulated cyclic nucleotide phosphodiesterase. Nature 459: 1015–1018 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous