

Semicircular canal morphogenesis in the zebrafish inner ear requires the function of gpr126 (lauscher), an adhesion class G protein-coupled receptor gene
- PMID: 24067352
- PMCID: PMC4007713
- DOI: 10.1242/dev.098061
Semicircular canal morphogenesis in the zebrafish inner ear requires the function of gpr126 (lauscher), an adhesion class G protein-coupled receptor gene
Abstract
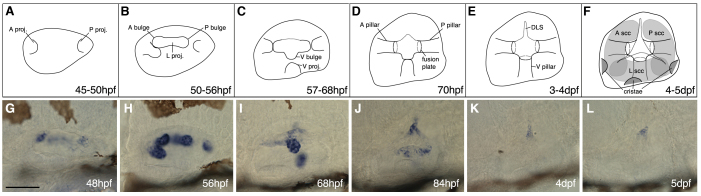
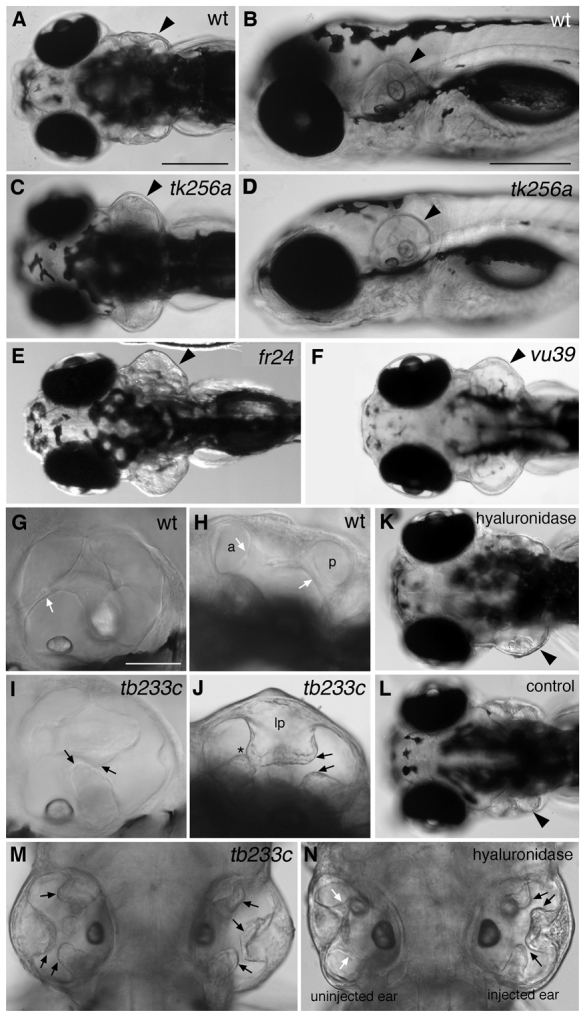
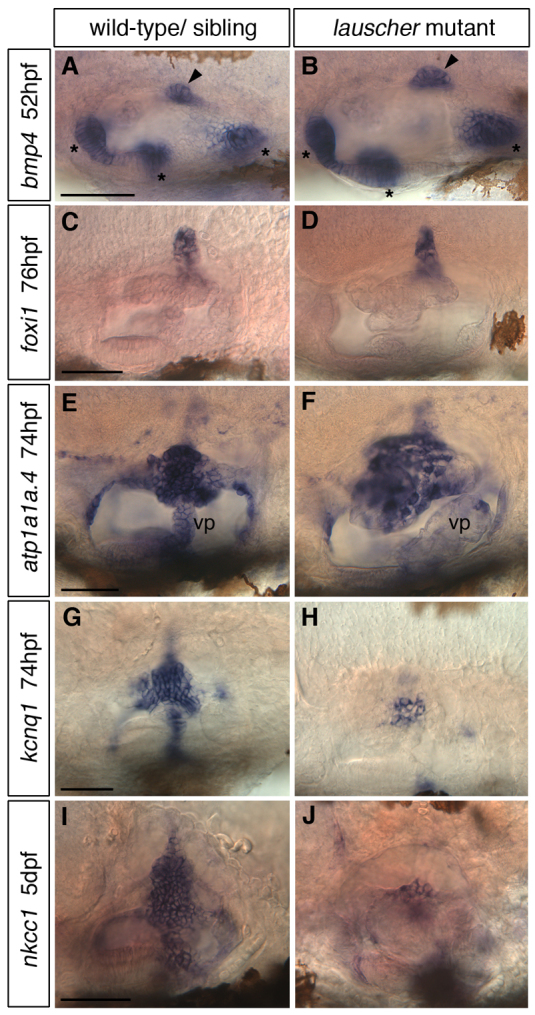
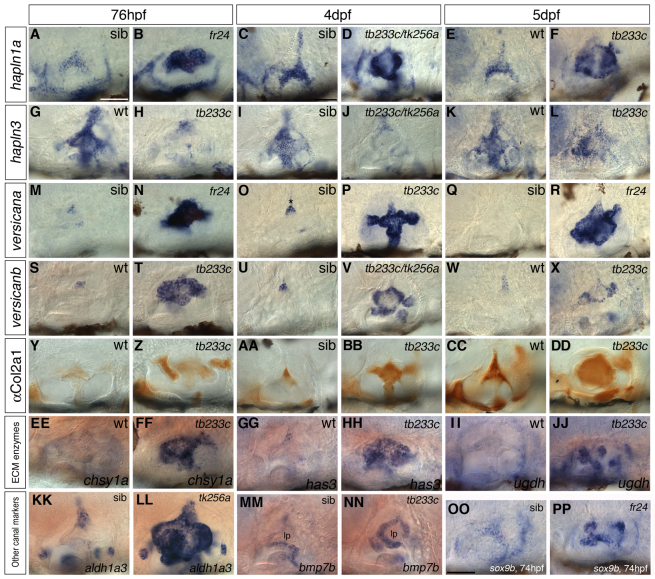
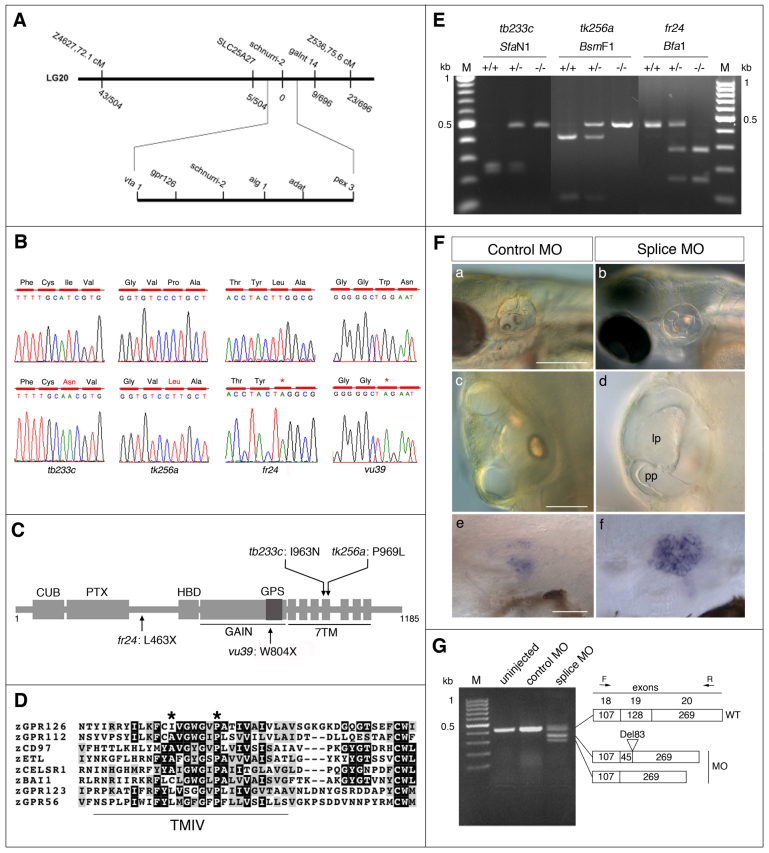
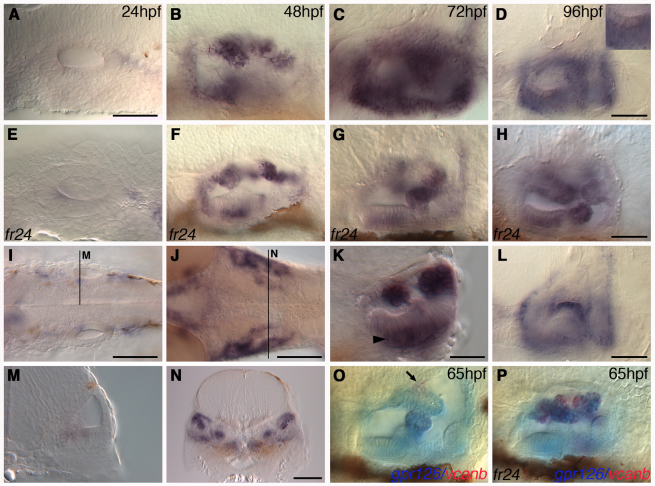
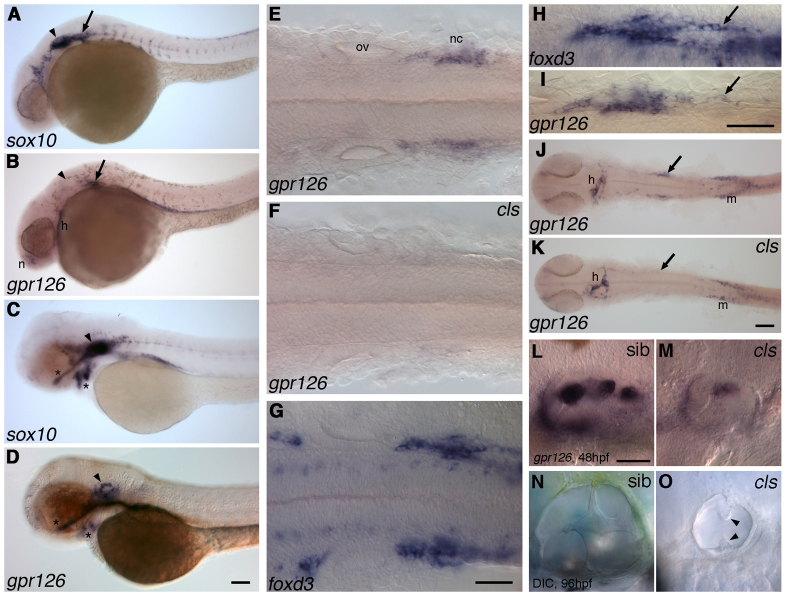
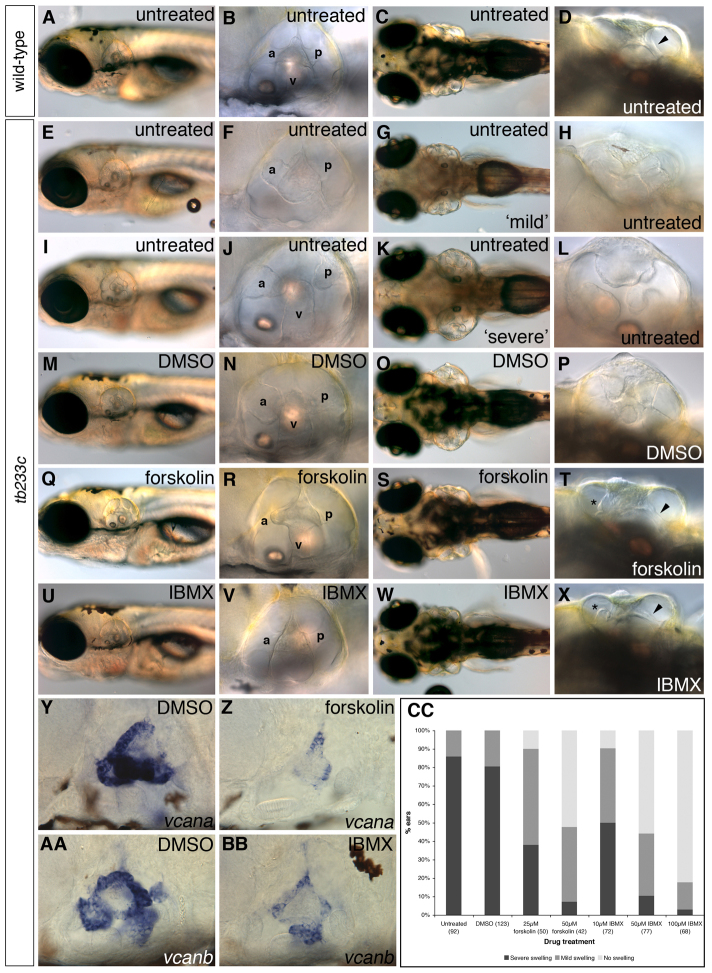
Morphogenesis of the semicircular canal ducts in the vertebrate inner ear is a dramatic example of epithelial remodelling in the embryo, and failure of normal canal development results in vestibular dysfunction. In zebrafish and Xenopus, semicircular canal ducts develop when projections of epithelium, driven by extracellular matrix production, push into the otic vesicle and fuse to form pillars. We show that in the zebrafish, extracellular matrix gene expression is high during projection outgrowth and then rapidly downregulated after fusion. Enzymatic disruption of hyaluronan in the projections leads to their collapse and a failure to form pillars: as a result, the ears swell. We have cloned a zebrafish mutant, lauscher (lau), identified by its swollen ear phenotype. The primary defect in the ear is abnormal projection outgrowth and a failure of fusion to form the semicircular canal pillars. Otic expression of extracellular matrix components is highly disrupted: several genes fail to become downregulated and remain expressed at abnormally high levels into late larval stages. The lau mutations disrupt gpr126, an adhesion class G protein-coupled receptor gene. Expression of gpr126 is similar to that of sox10, an ear and neural crest marker, and is partially dependent on sox10 activity. Fusion of canal projections and downregulation of otic versican expression in a hypomorphic lau allele can be restored by cAMP agonists. We propose that Gpr126 acts through a cAMP-mediated pathway to control the outgrowth and adhesion of canal projections in the zebrafish ear via the regulation of extracellular matrix gene expression.
Keywords: Adhesion GPCRs; Extracellular matrix; Inner ear; Semicircular canals; Zebrafish; gpr126.
Figures
References
-
- Acampora D., Merlo G. R., Paleari L., Zerega B., Postiglione M. P., Mantero S., Bober E., Barbieri O., Simeone A., Levi G. (1999). Craniofacial, vestibular and bone defects in mice lacking the Distal-less-related gene Dlx5. Development 126, 3795–3809 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
