

Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle
- PMID: 24067916
- PMCID: PMC3882385
- DOI: 10.1152/ajpcell.00171.2013
Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle
Abstract
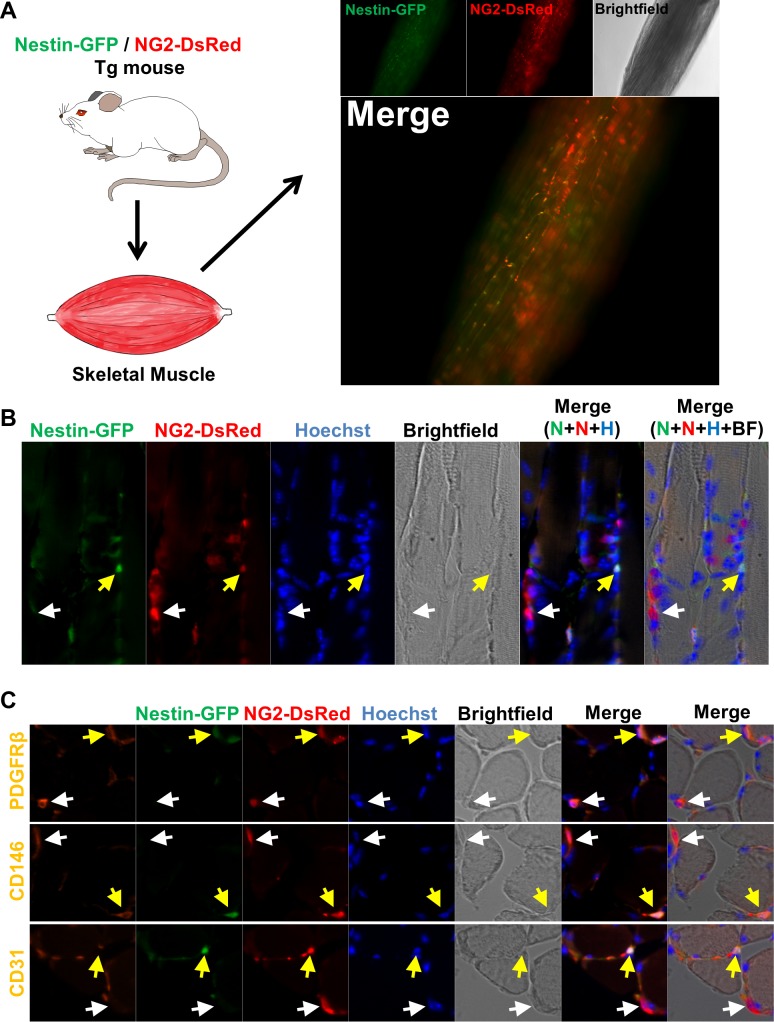
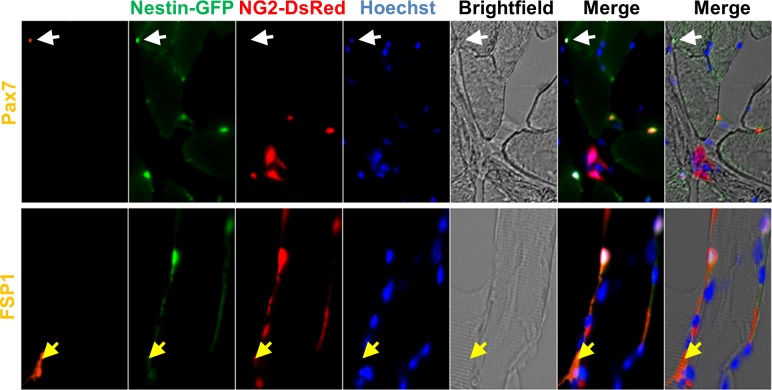
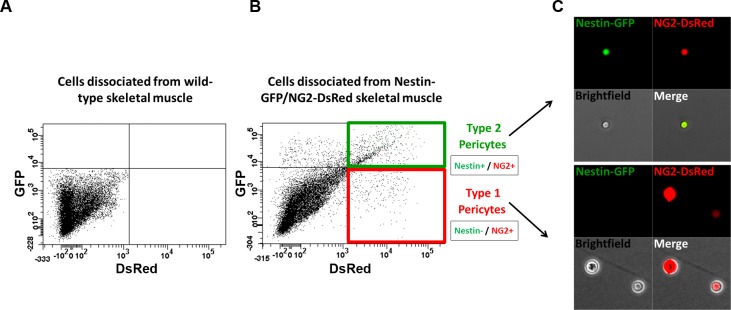
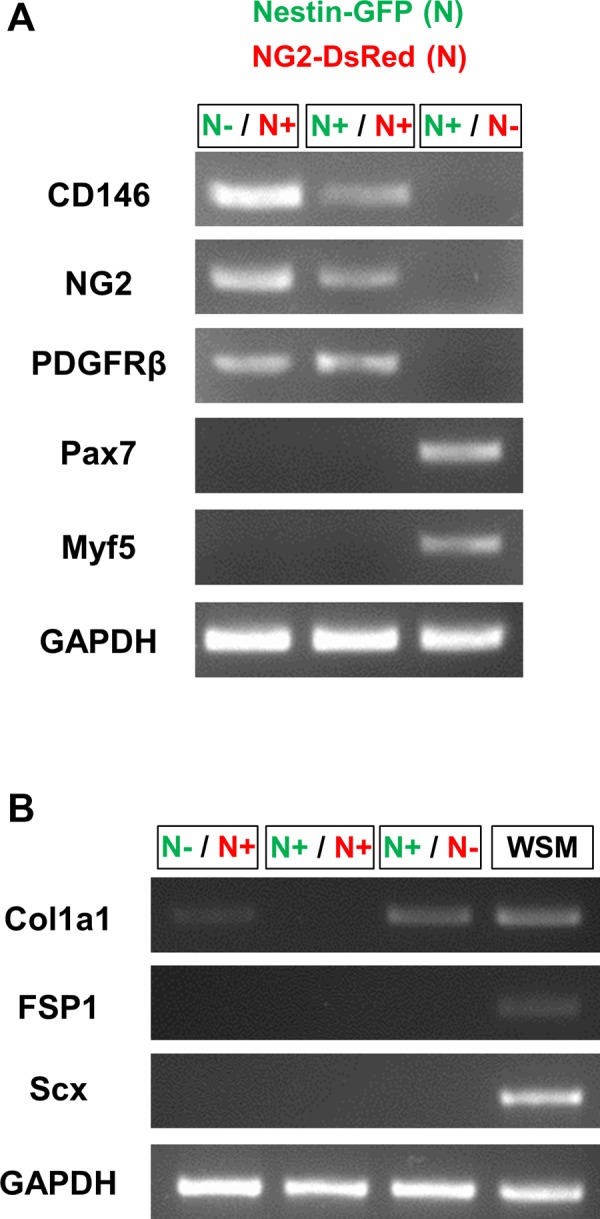
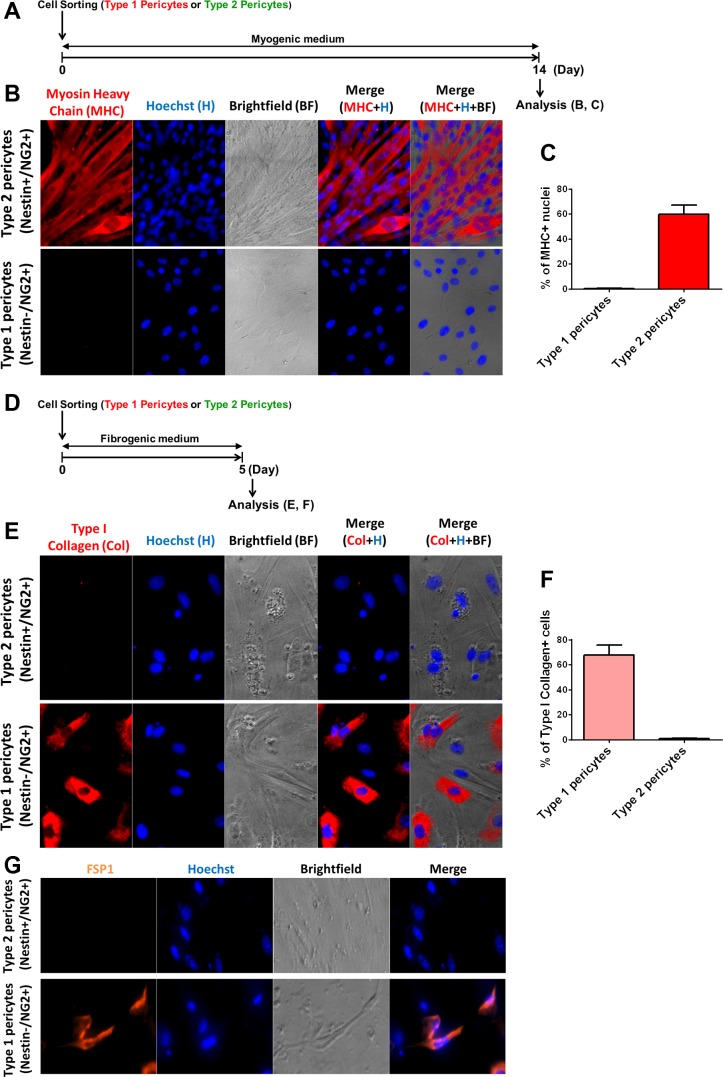
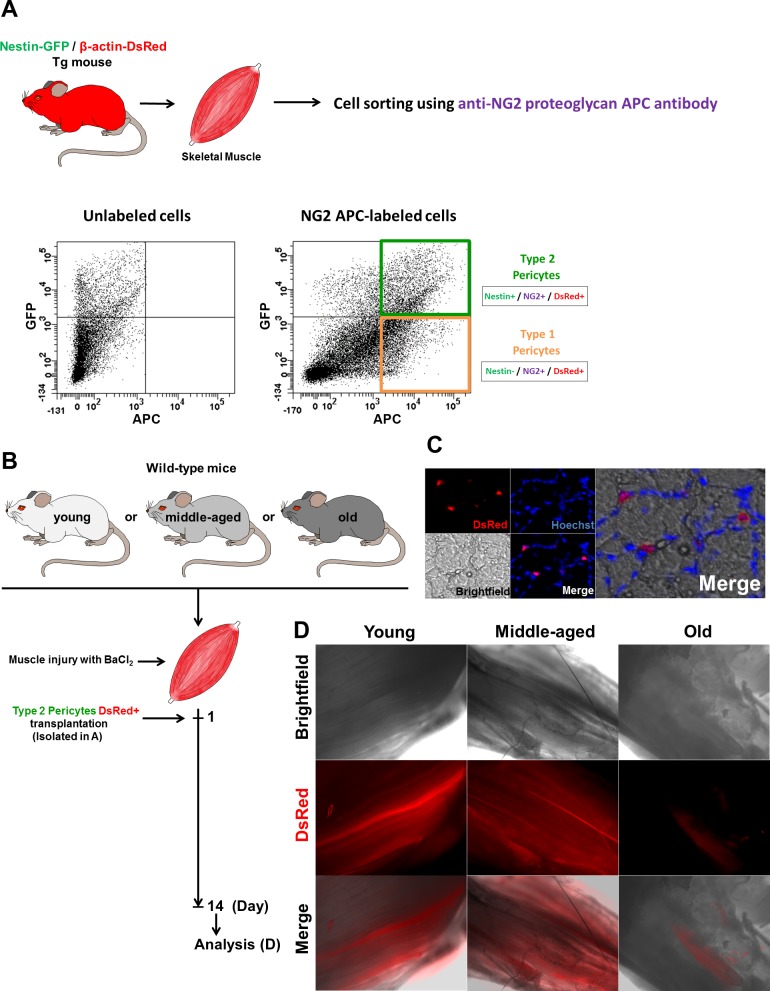
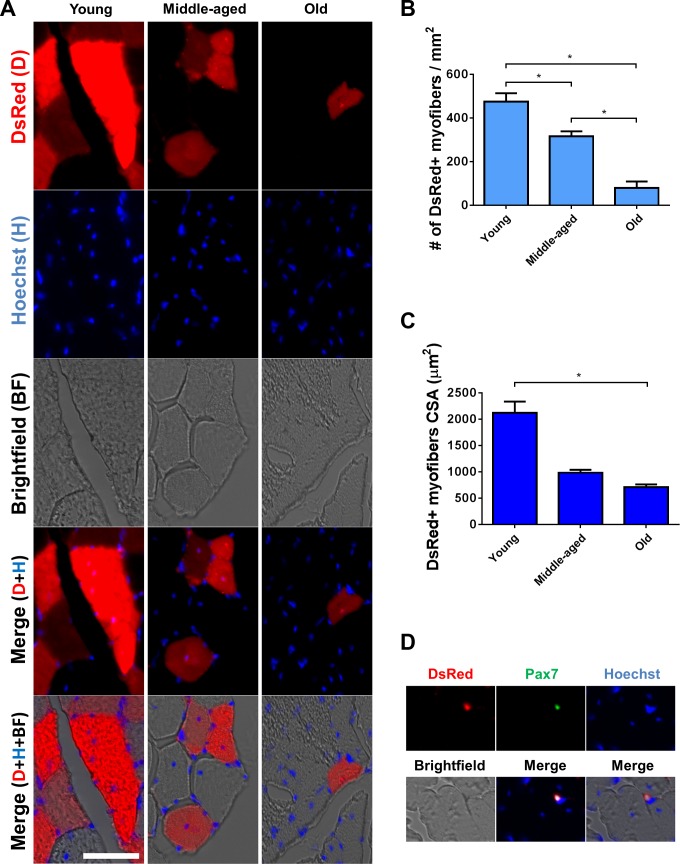
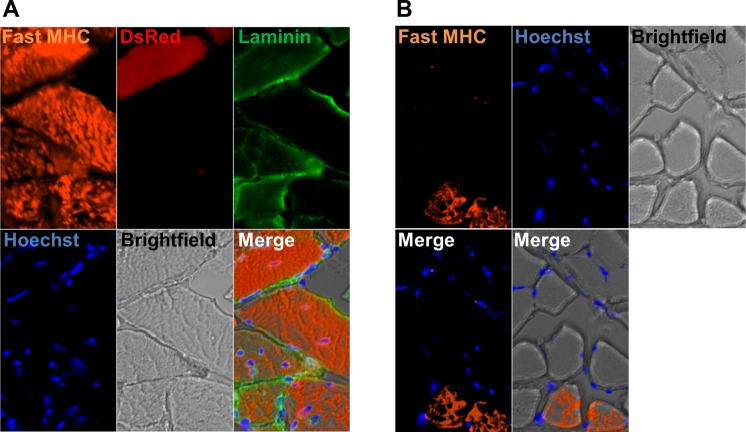
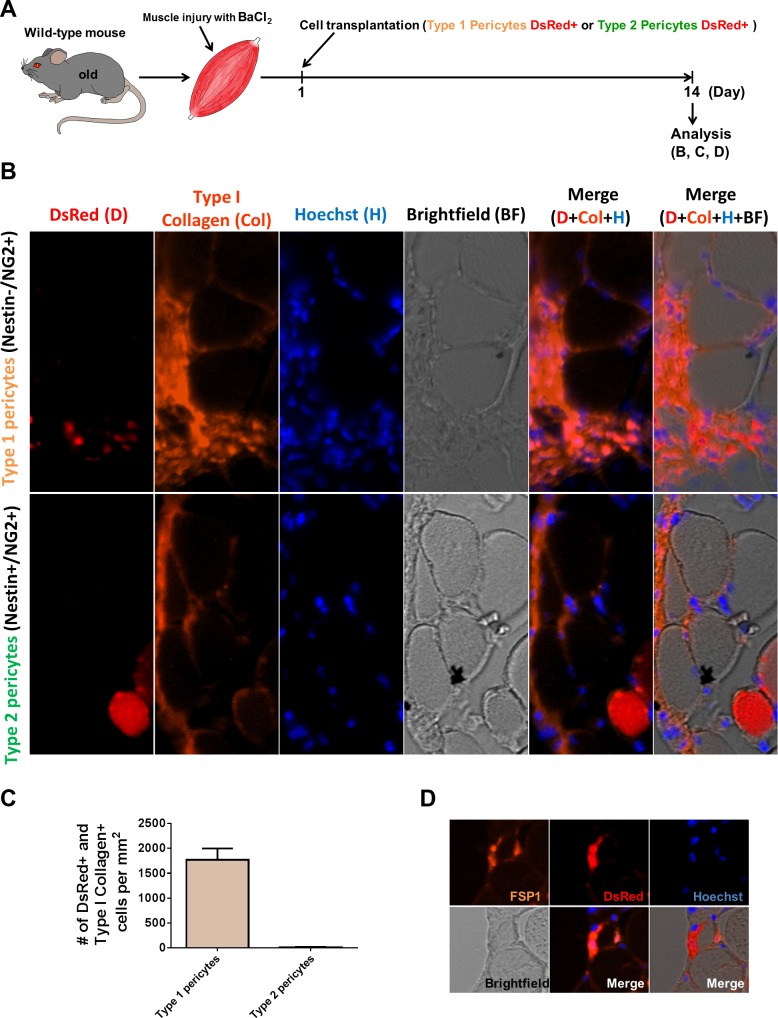
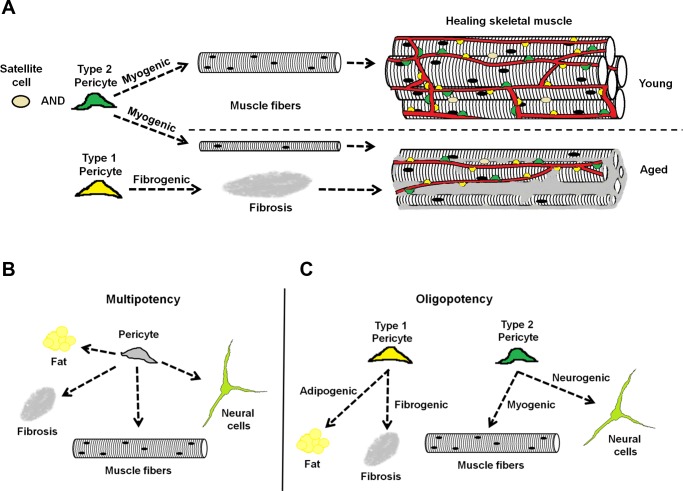
In older adults, changes in skeletal muscle composition are associated with increased fibrosis, loss of mass, and decreased force, which can lead to dependency, morbidity, and mortality. Understanding the biological mechanisms responsible is essential to sustaining and improving their quality of life. Compared with young mice, aged mice take longer to recover from muscle injury; their tissue fibrosis is more extensive, and regenerated myofibers are smaller. Strong evidence indicates that cells called pericytes, embedded in the basement membrane of capillaries, contribute to the satellite-cell pool and muscle growth. In addition to their role in skeletal muscle repair, after tissue damage, they detach from capillaries and migrate to the interstitial space to participate in fibrosis formation. Here we distinguish two bona fide pericyte subtypes in the skeletal muscle interstitium, type-1 (Nestin-GFP(-)/NG2-DsRed(+)) and type-2 (Nestin-GFP(+)/NG2-DsRed(+)), and characterize their heretofore unknown specific roles in the aging environment. Our in vitro results show that type-1 and type-2 pericytes are either fibrogenic or myogenic, respectively. Transplantation studies in young animals indicate that type-2 pericytes are myogenic, while type-1 pericytes remain in the interstitial space. In older mice, however, the muscular regenerative capacity of type-2 pericytes is limited, and type-1 pericytes produce collagen, contributing to fibrous tissue deposition. We conclude that in injured muscles from aging mice, the pericytes involved in skeletal muscle repair differ from those associated with scar formation.
Keywords: aging; fibrous tissue; pericytes; skeletal muscle.
Figures
References
-
- Barnes JL, Glass WF., 2nd Renal interstitial fibrosis: a critical evaluation of the origin of myofibroblasts. Contrib Nephrol 169: 73–93, 2011 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases