

Comparative analysis of adeno-associated virus capsid stability and dynamics
- PMID: 24067976
- PMCID: PMC3838259
- DOI: 10.1128/JVI.01415-13
Comparative analysis of adeno-associated virus capsid stability and dynamics
Abstract
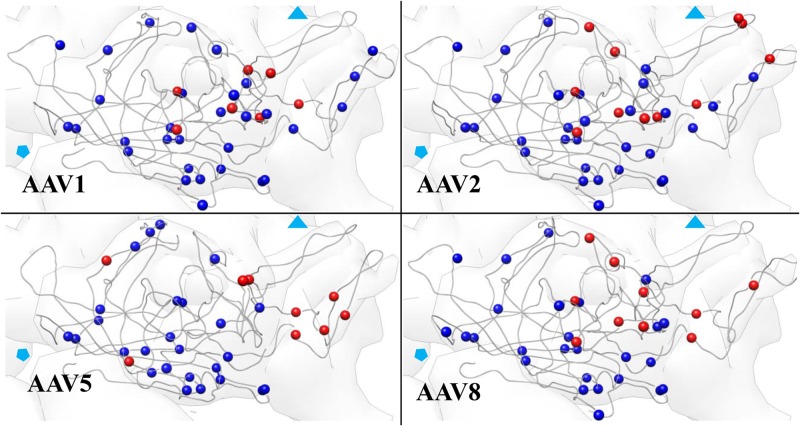
Icosahedral viral capsids are obligated to perform a thermodynamic balancing act. Capsids must be stable enough to protect the genome until a suitable host cell is encountered yet be poised to bind receptor, initiate cell entry, navigate the cellular milieu, and release their genome in the appropriate replication compartment. In this study, serotypes of adeno-associated virus (AAV), AAV1, AAV2, AAV5, and AAV8, were compared with respect to the physical properties of their capsids that influence thermodynamic stability. Thermal stability measurements using differential scanning fluorimetry, differential scanning calorimetry, and electron microscopy showed that capsid melting temperatures differed by more than 20°C between the least and most stable serotypes, AAV2 and AAV5, respectively. Limited proteolysis and peptide mass mapping of intact particles were used to investigate capsid protein dynamics. Active hot spots mapped to the region surrounding the 3-fold axis of symmetry for all serotypes. Cleavages also mapped to the unique region of VP1 which contains a phospholipase domain, indicating transient exposure on the surface of the capsid. Data on the biophysical properties of the different AAV serotypes are important for understanding cellular trafficking and is critical to their production, storage, and use for gene therapy. The distinct differences reported here provide direction for future studies on entry and vector production.
Figures






References
-
- Halder S, Ng R, Agbandje-McKenna M. 2012. Parvoviruses: structure and infection. Future Virol. 7:253–278
-
- Muzyczka N, Berns KI. 2001. Parvoviridae: the viruses and their replication, p 2327–2360 In Knipe DM, Howley PM. (ed), Fields virology, 4th ed. Lippincott Williams and Wilkins, New York, NY
-
- Agbandje-McKenna M, Kleinschmidt J. 2011. AAV capsid structure and cell interactions. Methods Mol. Biol. 807:47–92 - PubMed
-
- Agbandje-McKenna M, Chapman MS. 2006. Correlating structure with function in the viral capsid, p 125–139 In Kerr JR, Cotmore SF, Bloom ME, Linden R, Parrish CR. (ed), Parvoviruses. Hodder Arnold Publication, London, United Kingdom
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
