

Atypical E2fs control lymphangiogenesis through transcriptional regulation of Ccbe1 and Flt4
- PMID: 24069224
- PMCID: PMC3771987
- DOI: 10.1371/journal.pone.0073693
Atypical E2fs control lymphangiogenesis through transcriptional regulation of Ccbe1 and Flt4
Erratum in
- PLoS One. 2013;8(9). doi:10.1371/annotation/25c7c486-9e68-4df9-af01-2fe1c5d1a2d4
Abstract
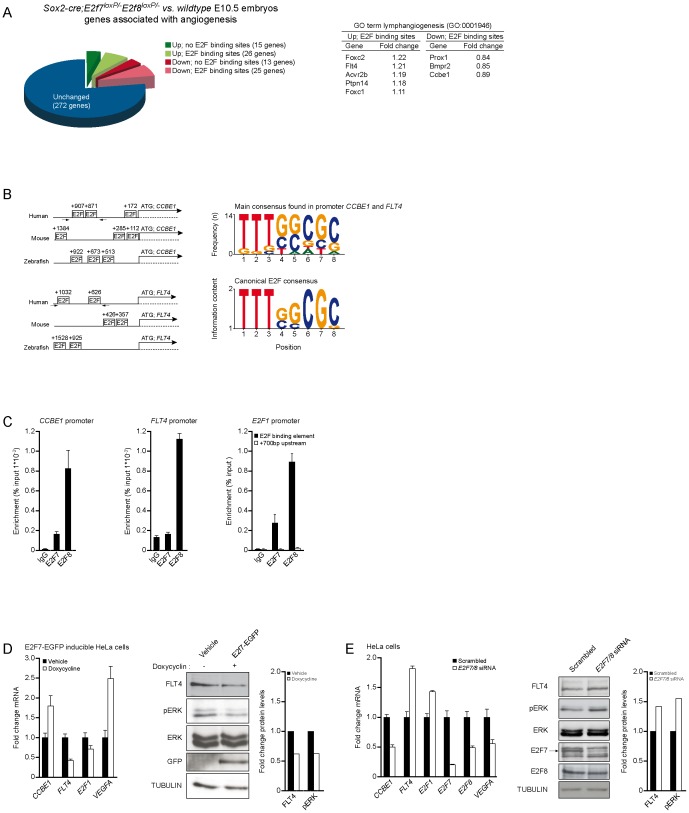
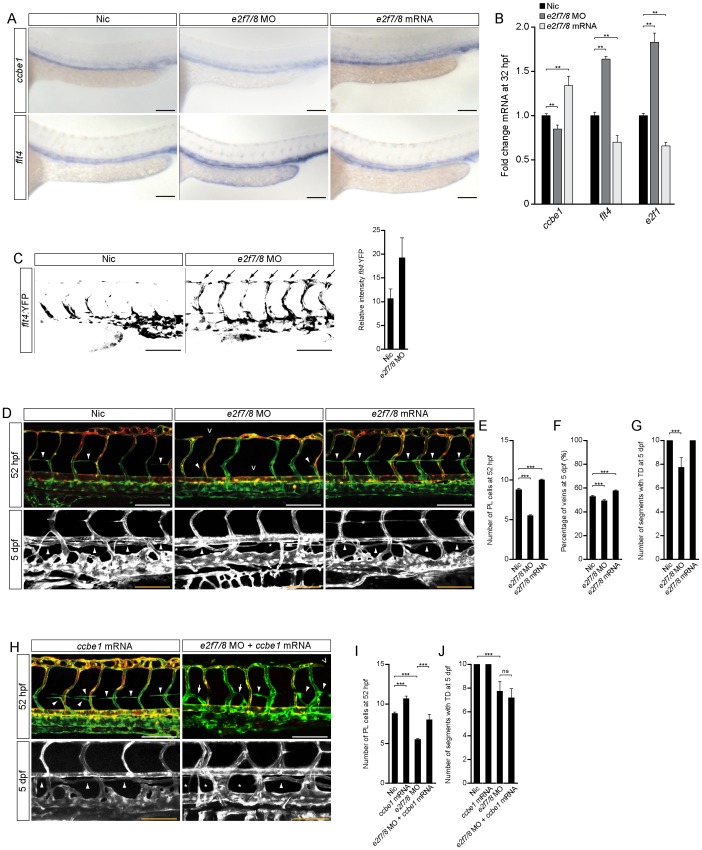
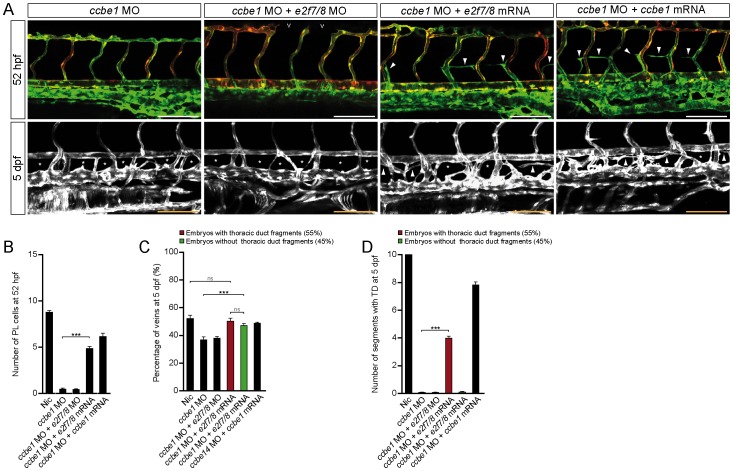
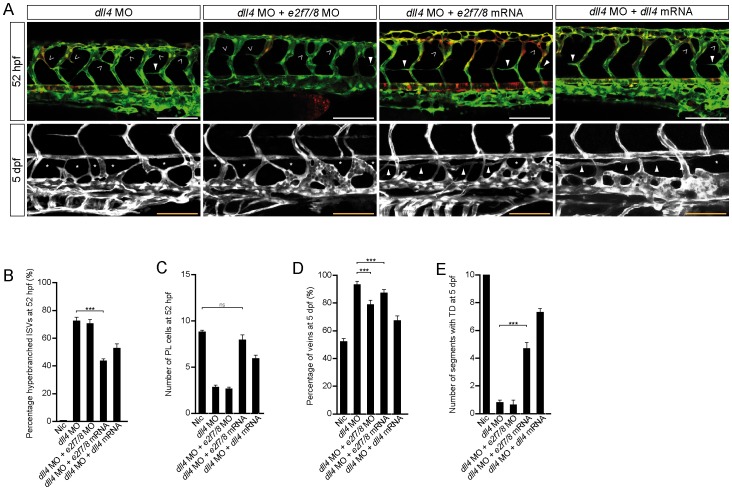
Lymphatic vessels are derived from venous endothelial cells and their formation is governed by the Vascular endothelial growth factor C (VegfC)/Vegf receptor 3 (Vegfr3; Flt4) signaling pathway. Recent studies show that Collagen and Calcium Binding EGF domains 1 protein (Ccbe1) enhances VegfC-dependent lymphangiogenesis. Both Ccbe1 and Flt4 have been shown to be indispensable for lymphangiogenesis. However, how these essential players are transcriptionally regulated remains poorly understood. In the case of angiogenesis, atypical E2fs (E2f7 and E2f8) however have been recently shown to function as transcriptional activators for VegfA. Using a genome-wide approach we here identified both CCBE1 and FLT4 as direct targets of atypical E2Fs. E2F7/8 directly bind and stimulate the CCBE1 promoter, while recruitment of E2F7/8 inhibits the FLT4 promoter. Importantly, inactivation of e2f7/8 in zebrafish impaired venous sprouting and lymphangiogenesis with reduced ccbe1 expression and increased flt4 expression. Remarkably, over-expression of e2f7/8 rescued Ccbe1- and Flt4-dependent lymphangiogenesis phenotypes. Together these results identified E2f7/8 as novel in vivo transcriptional regulators of Ccbe1 and Flt4, both essential genes for venous sprouting and lymphangiogenesis.
Conflict of interest statement
Figures
Similar articles
-
Ccbe1 regulates Vegfc-mediated induction of Vegfr3 signaling during embryonic lymphangiogenesis.Development. 2014 Mar;141(6):1239-49. doi: 10.1242/dev.100495. Epub 2014 Feb 12. Development. 2014. PMID: 24523457
-
ETS transcription factor Etsrp / Etv2 is required for lymphangiogenesis and directly regulates vegfr3 / flt4 expression.Dev Biol. 2018 Aug 1;440(1):40-52. doi: 10.1016/j.ydbio.2018.05.003. Epub 2018 May 9. Dev Biol. 2018. PMID: 29753018 Free PMC article.
-
CCBE1 enhances lymphangiogenesis via A disintegrin and metalloprotease with thrombospondin motifs-3-mediated vascular endothelial growth factor-C activation.Circulation. 2014 May 13;129(19):1962-71. doi: 10.1161/CIRCULATIONAHA.113.002779. Epub 2014 Feb 19. Circulation. 2014. PMID: 24552833
-
A fisheye view on lymphangiogenesis.Adv Anat Embryol Cell Biol. 2014;214:153-65. doi: 10.1007/978-3-7091-1646-3_12. Adv Anat Embryol Cell Biol. 2014. PMID: 24276893 Review.
-
The lymphatic vasculature revisited-new developments in the zebrafish.Methods Cell Biol. 2017;138:221-238. doi: 10.1016/bs.mcb.2016.11.001. Epub 2016 Dec 29. Methods Cell Biol. 2017. PMID: 28129845 Review.
Cited by
-
Harnessing Gene Expression Profiles for the Identification of Ex Vivo Drug Response Genes in Pediatric Acute Myeloid Leukemia.Cancers (Basel). 2020 May 15;12(5):1247. doi: 10.3390/cancers12051247. Cancers (Basel). 2020. PMID: 32429253 Free PMC article.
-
A Multiplex Kindred with Hennekam Syndrome due to Homozygosity for a CCBE1 Mutation that does not Prevent Protein Expression.J Clin Immunol. 2016 Jan;36(1):19-27. doi: 10.1007/s10875-015-0225-6. Epub 2015 Dec 19. J Clin Immunol. 2016. PMID: 26686525 Free PMC article.
-
Upregulation of E2F8 promotes cell proliferation and tumorigenicity in breast cancer by modulating G1/S phase transition.Oncotarget. 2016 Apr 26;7(17):23757-71. doi: 10.18632/oncotarget.8121. Oncotarget. 2016. PMID: 26992224 Free PMC article.
-
Interactions between genes altered during cardiotoxicity and neurotoxicity in zebrafish revealed using induced network modules analysis.Sci Rep. 2023 Apr 17;13(1):6257. doi: 10.1038/s41598-023-33145-8. Sci Rep. 2023. PMID: 37069190 Free PMC article.
-
Whole blood transcript and protein abundance of the vascular endothelial growth factor family relate to cognitive performance.Neurobiol Aging. 2023 Apr;124:11-17. doi: 10.1016/j.neurobiolaging.2023.01.002. Epub 2023 Jan 7. Neurobiol Aging. 2023. PMID: 36680854 Free PMC article.
References
-
- Tammela T, Alitalo K (2010) Lymphangiogenesis: Molecular mechanisms and future promise Cell. 140: 460–476. - PubMed
-
- Hogan BM, Herpers R, Witte M, Helotera H, Alitalo K, et al. (2009) Vegfc/Flt4 signalling is suppressed by Dll4 in developing zebrafish intersegmental arteries Development. 136: 4001–4009. - PubMed
-
- Karkkainen MJ, Haiko P, Sainio K, Partanen J, Taipale J, et al. (2004) Vascular endothelial growth factor C is required for sprouting of the first lymphatic vessels from embryonic veins Nat Immunol. 5: 74–80. - PubMed
-
- Bos FL, Caunt M, Peterson-Maduro J, Planas-Paz L, Kowalski J, et al. (2011) CCBE1 is essential for mammalian lymphatic vascular development and enhances the lymphangiogenic effect of vascular endothelial growth factor-C in vivo Circ Res. 109: 486–491. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
