

Combined inhibition of p97 and the proteasome causes lethal disruption of the secretory apparatus in multiple myeloma cells
- PMID: 24069311
- PMCID: PMC3775786
- DOI: 10.1371/journal.pone.0074415
Combined inhibition of p97 and the proteasome causes lethal disruption of the secretory apparatus in multiple myeloma cells
Abstract
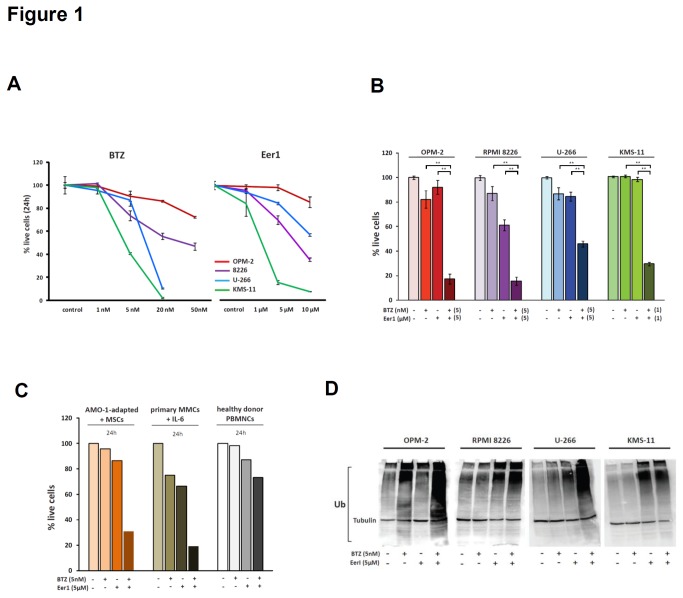
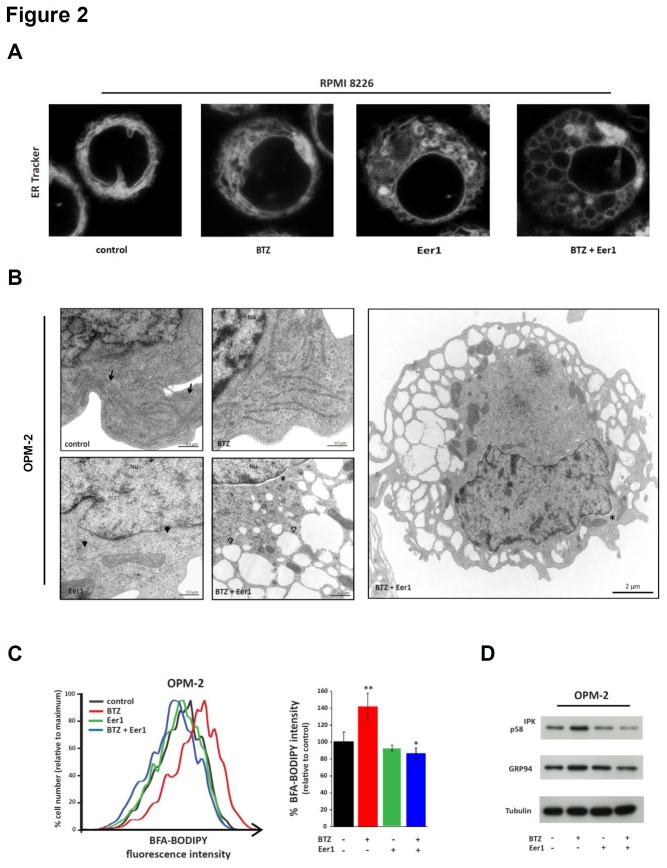
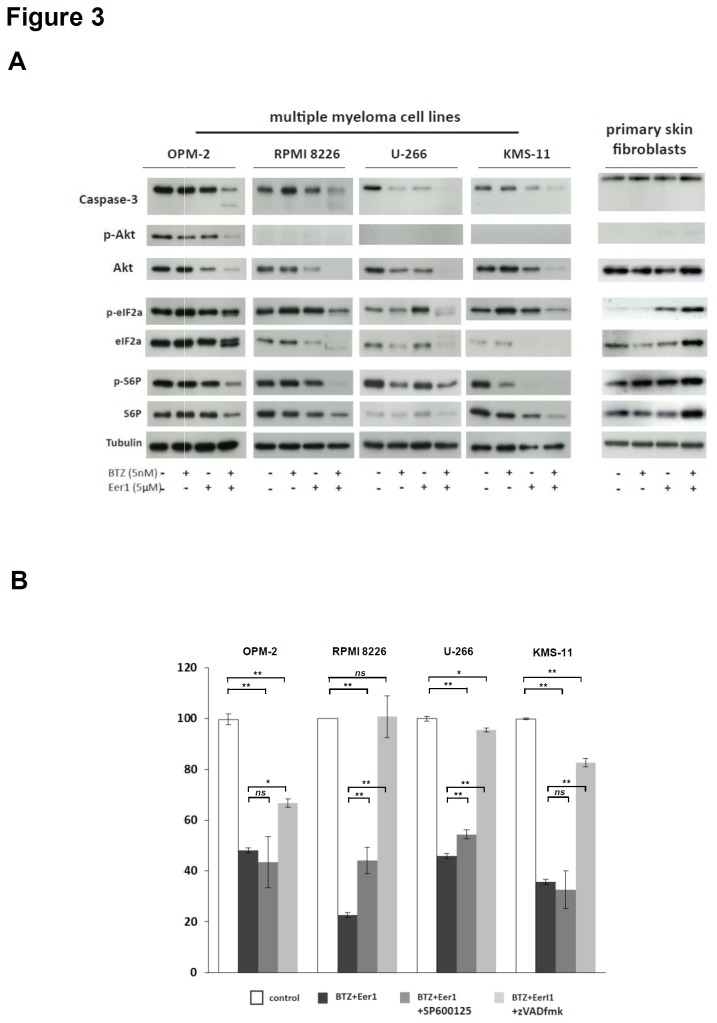
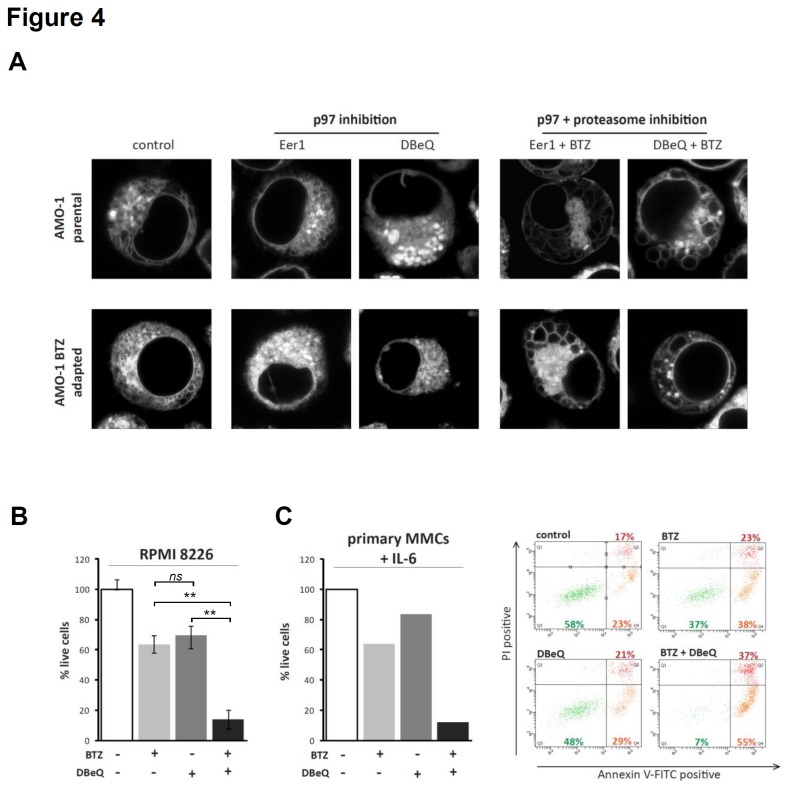
Inhibition of the proteasome is a widely used strategy for treating multiple myeloma that takes advantage of the heavy secretory load that multiple myeloma cells (MMCs) have to deal with. Resistance of MMCs to proteasome inhibition has been linked to incomplete disruption of proteasomal endoplasmic-reticulum (ER)-associated degradation (ERAD) and activation of non-proteasomal protein degradation pathways. The ATPase p97 (VCP/Cdc48) has key roles in mediating both ERAD and non-proteasomal protein degradation and can be targeted pharmacologically by small molecule inhibition. In this study, we compared the effects of p97 inhibition with Eeyarestatin 1 and DBeQ on the secretory apparatus of MMCs with the effects induced by the proteasome inhibitor bortezomib, and the effects caused by combined inhibition of p97 and the proteasome. We found that p97 inhibition elicits cellular responses that are different from those induced by proteasome inhibition, and that the responses differ considerably between MMC lines. Moreover, we found that dual inhibition of both p97 and the proteasome terminally disrupts ER configuration and intracellular protein metabolism in MMCs. Dual inhibition of p97 and the proteasome induced high levels of apoptosis in all of the MMC lines that we analysed, including bortezomib-adapted AMO-1 cells, and was also effective in killing primary MMCs. Only minor toxicity was observed in untransformed and non-secretory cells. Our observations highlight non-redundant roles of p97 and the proteasome in maintaining secretory homeostasis in MMCs and provide a preclinical conceptual framework for dual targeting of p97 and the proteasome as a potential new therapeutic strategy in multiple myeloma.
Conflict of interest statement
Figures
References
-
- Aronson LI, Davies FE (2012) DangER: protein ovERload. Targeting protein degradation to treat myeloma. Haematologica 97: 1119-1130. doi:10.3324/haematol.2012.064923. PubMed: 22580998. - DOI - PMC - PubMed
-
- Schröder M, Kaufman RJ (2005) The mammalian unfolded protein response. Annu Rev Biochem 74: 739-789. doi:10.1146/annurev.biochem.73.011303.074134. PubMed: 15952902. - DOI - PubMed
-
- Vembar SS, Brodsky JL (2008) One step at a time: endoplasmic reticulum-associated degradation. Nat Rev Mol Cell Biol 9: 944-957. doi:10.1038/nrm2546. PubMed: 19002207. - DOI - PMC - PubMed
-
- Tabas I, Ron D (2011) Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat Cell Biol 13: 184-190. doi:10.1038/ncb0311-184. PubMed: 21364565. - DOI - PMC - PubMed
-
- Bianchi G, Oliva L, Cascio P, Pengo N, Fontana F et al. (2009) The proteasome load versus capacity balance determines apoptotic sensitivity of multiple myeloma cells to proteasome inhibition. Blood 113: 3040-3049. doi:10.1182/blood-2008-08-172734. PubMed: 19164601. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
