

Mu opioid receptors on primary afferent nav1.8 neurons contribute to opiate-induced analgesia: insight from conditional knockout mice
- PMID: 24069332
- PMCID: PMC3771900
- DOI: 10.1371/journal.pone.0074706
Mu opioid receptors on primary afferent nav1.8 neurons contribute to opiate-induced analgesia: insight from conditional knockout mice
Abstract
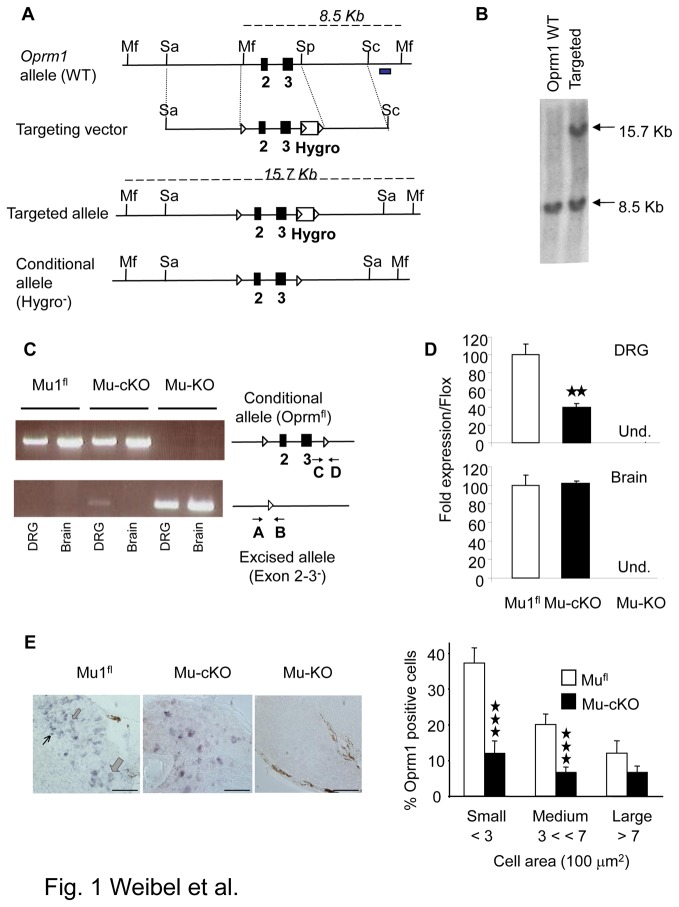
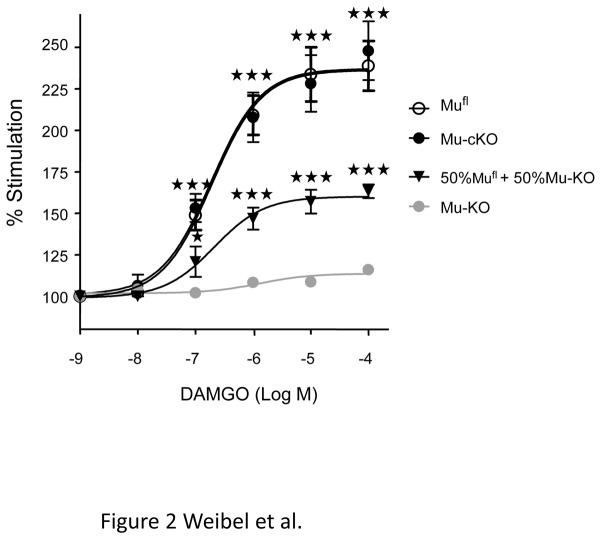
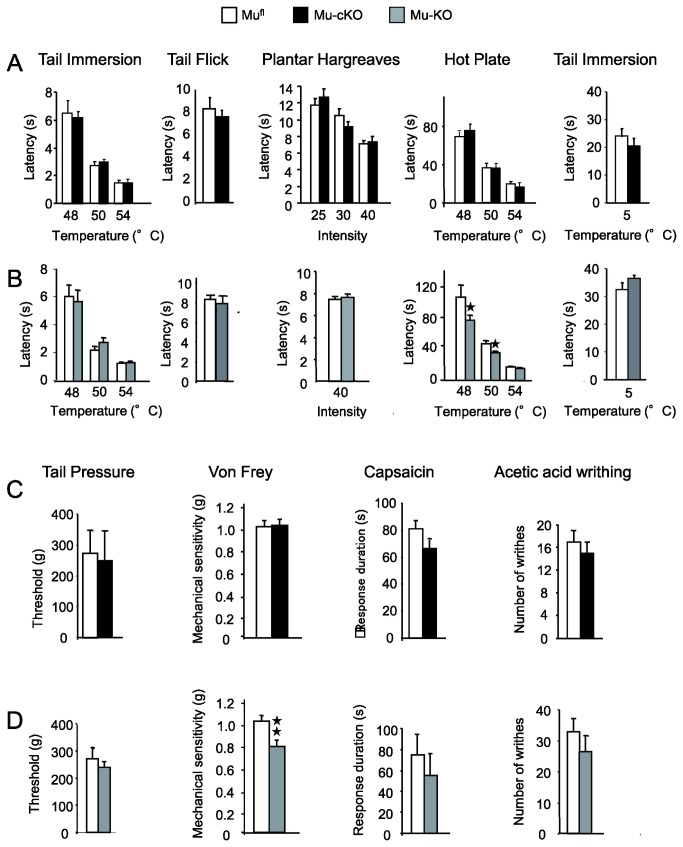
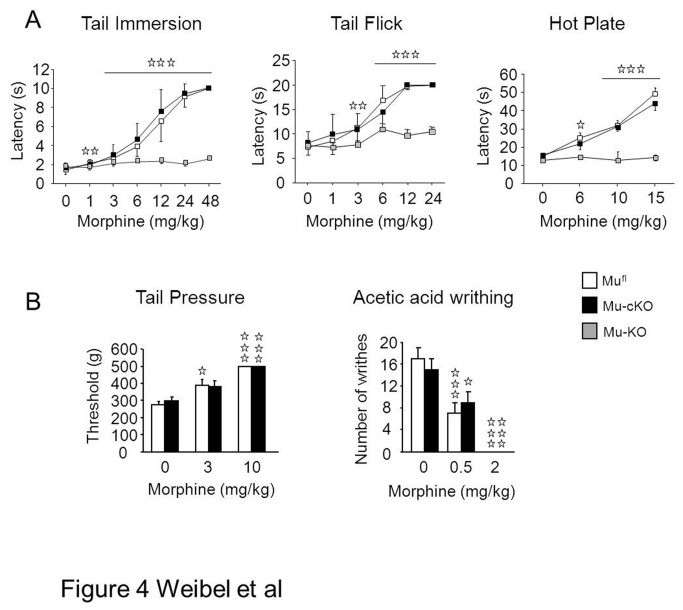
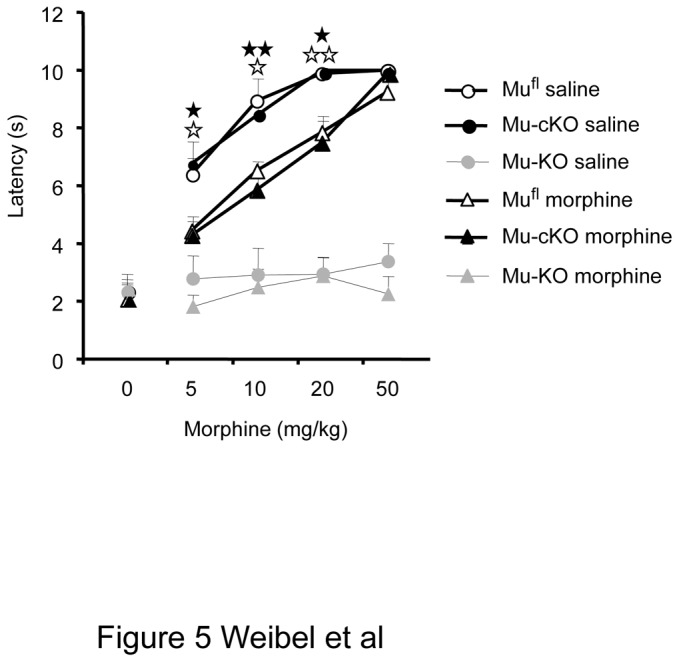
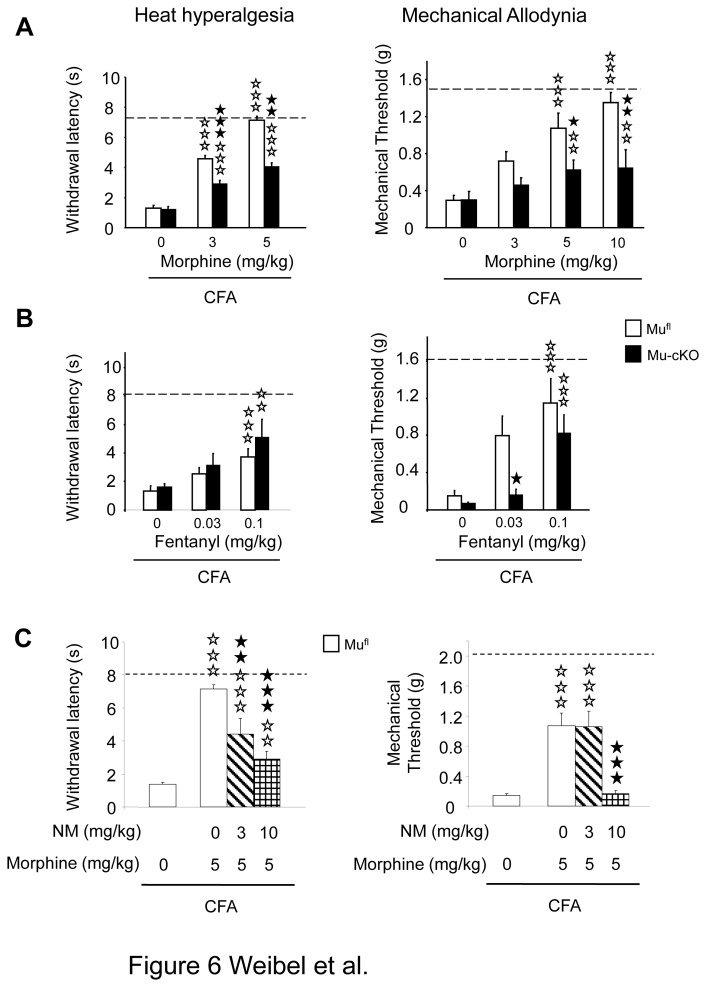
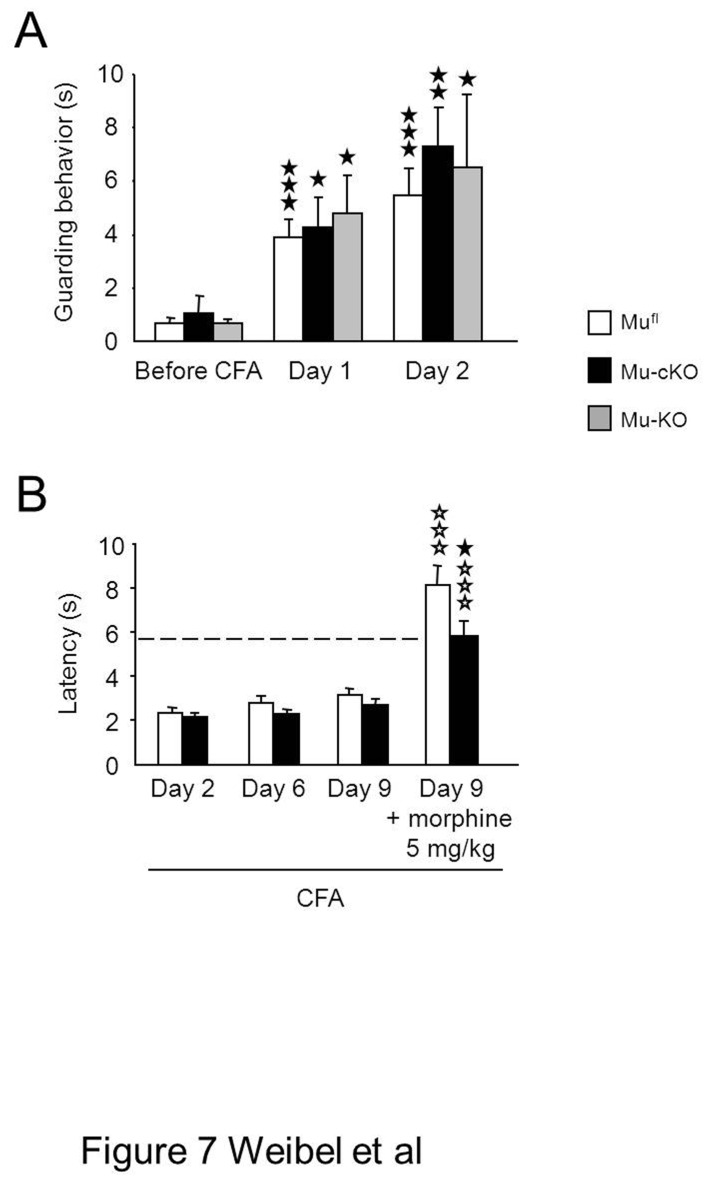
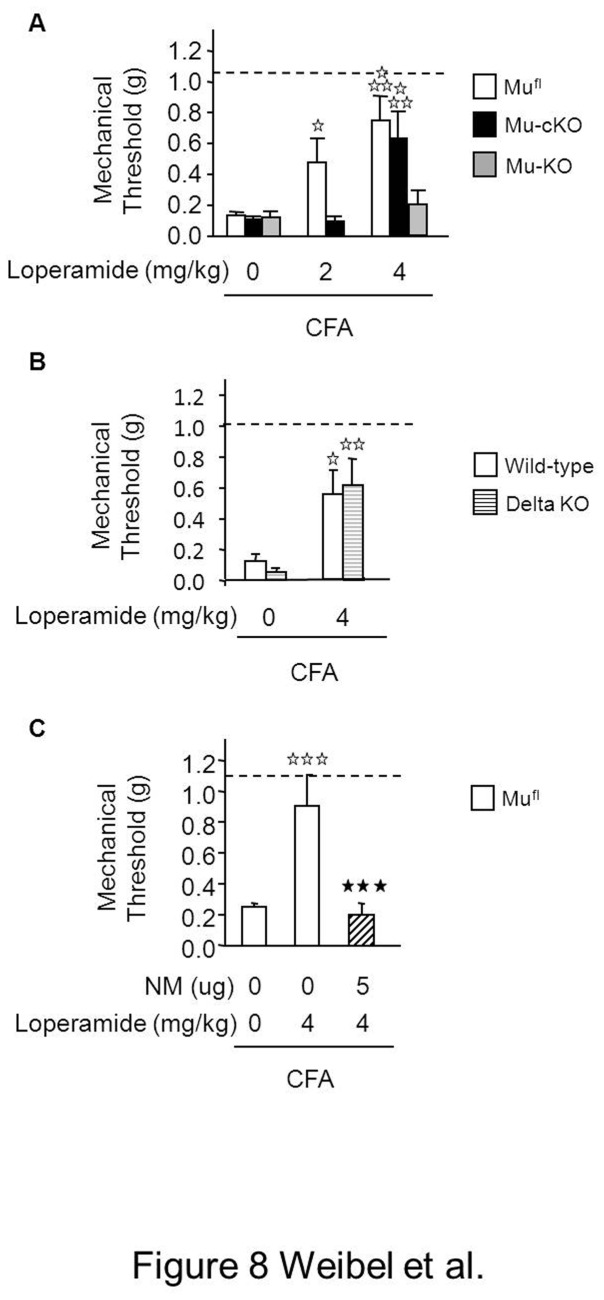
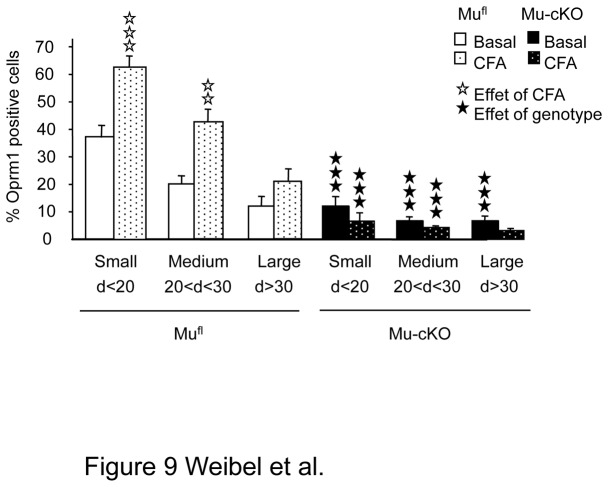
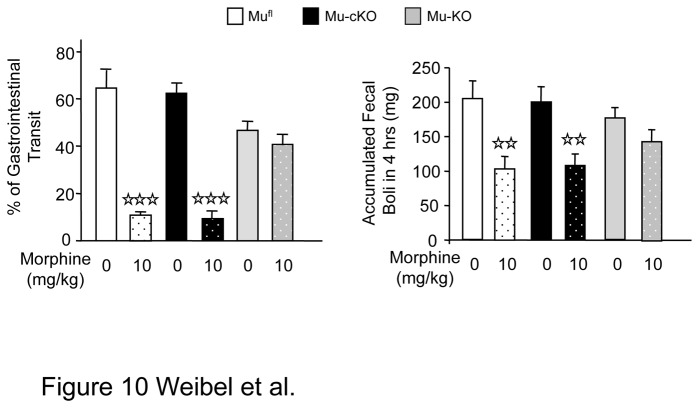
Opiates are powerful drugs to treat severe pain, and act via mu opioid receptors distributed throughout the nervous system. Their clinical use is hampered by centrally-mediated adverse effects, including nausea or respiratory depression. Here we used a genetic approach to investigate the potential of peripheral mu opioid receptors as targets for pain treatment. We generated conditional knockout (cKO) mice in which mu opioid receptors are deleted specifically in primary afferent Nav1.8-positive neurons. Mutant animals were compared to controls for acute nociception, inflammatory pain, opiate-induced analgesia and constipation. There was a 76% decrease of mu receptor-positive neurons and a 60% reduction of mu-receptor mRNA in dorsal root ganglia of cKO mice. Mutant mice showed normal responses to heat, mechanical, visceral and chemical stimuli, as well as unchanged morphine antinociception and tolerance to antinociception in models of acute pain. Inflammatory pain developed similarly in cKO and controls mice after Complete Freund's Adjuvant. In the inflammation model, however, opiate-induced (morphine, fentanyl and loperamide) analgesia was reduced in mutant mice as compared to controls, and abolished at low doses. Morphine-induced constipation remained intact in cKO mice. We therefore genetically demonstrate for the first time that mu opioid receptors partly mediate opiate analgesia at the level of Nav1.8-positive sensory neurons. In our study, this mechanism operates under conditions of inflammatory pain, but not nociception. Previous pharmacology suggests that peripheral opiates may be clinically useful, and our data further demonstrate that Nav1.8 neuron-associated mu opioid receptors are feasible targets to alleviate some forms of persistent pain.
Conflict of interest statement
Figures
References
-
- Le Merrer J, Becker JA, Befort K, Kieffer BL (2009) Reward processing by the opioid system in the brain. Physiol Rev 89: 1379-1412. doi:10.1152/physrev.00005.2009. PubMed: 19789384. - DOI - PMC - PubMed
-
- Mansour A, Fox CA, Akil H, Watson SJ (1995) Opioid-receptor mRNA expression in the rat CNS: anatomical and functional implications. Trends Neurosci 18: 22-29. doi:10.1016/0166-2236(95)93946-U. PubMed: 7535487. - DOI - PubMed
-
- Stein C, Machelska H (2011) Modulation of peripheral sensory neurons by the immune system: implications for pain therapy. Pharmacol Rev 63: 860-881. doi:10.1124/pr.110.003145. PubMed: 21969325. - DOI - PubMed
-
- Ossipov MH, Dussor GO, Porreca F (2010) Central modulation of pain. J Clin Invest 120: 3779-3787. doi:10.1172/JCI43766. PubMed: 21041960. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
