

Comparison of phylogeny, venom composition and neutralization by antivenom in diverse species of bothrops complex
- PMID: 24069493
- PMCID: PMC3772048
- DOI: 10.1371/journal.pntd.0002442
Comparison of phylogeny, venom composition and neutralization by antivenom in diverse species of bothrops complex
Abstract
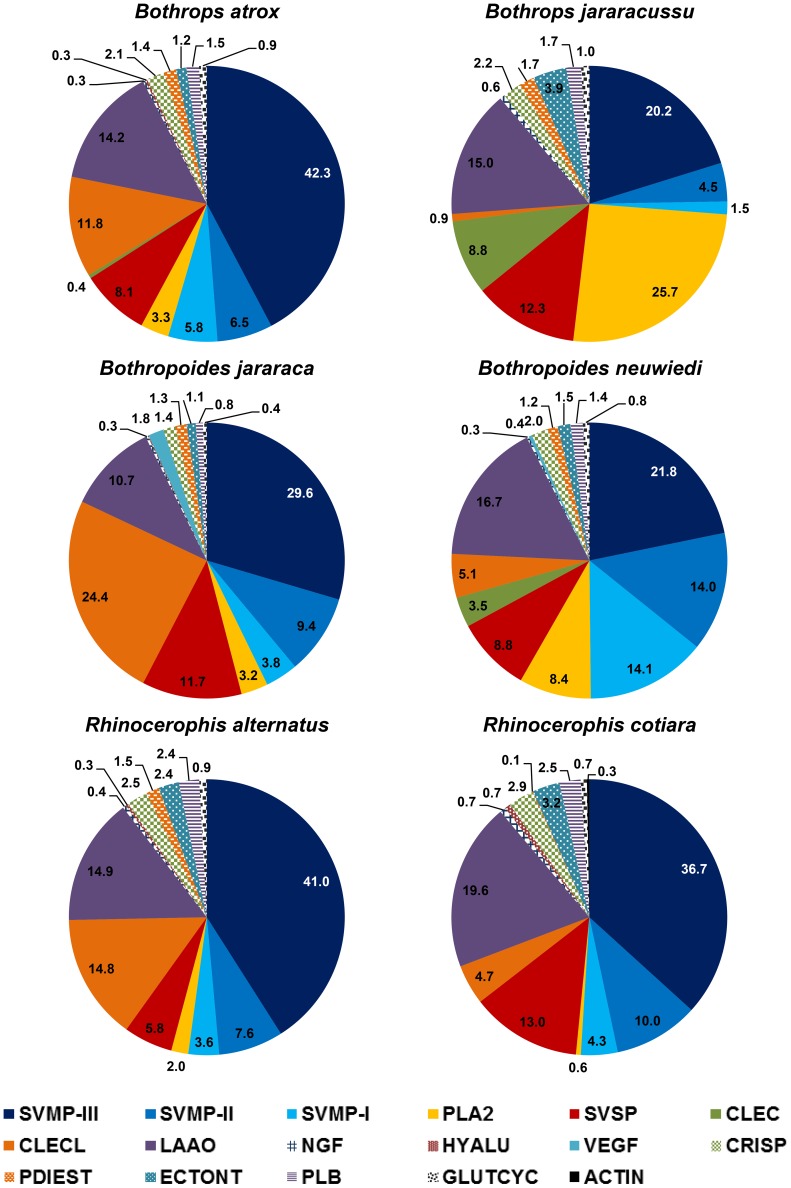
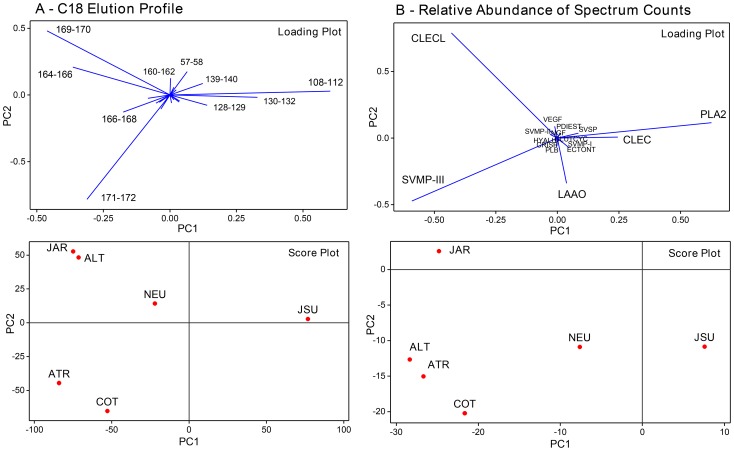
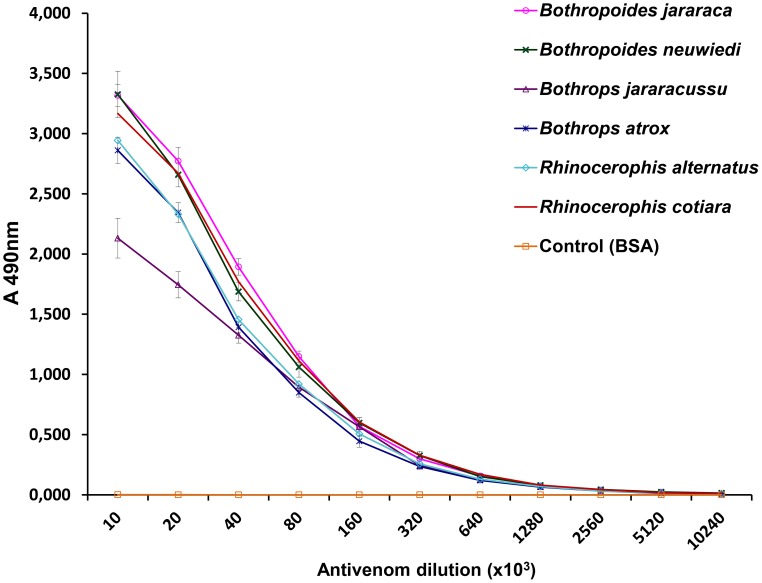
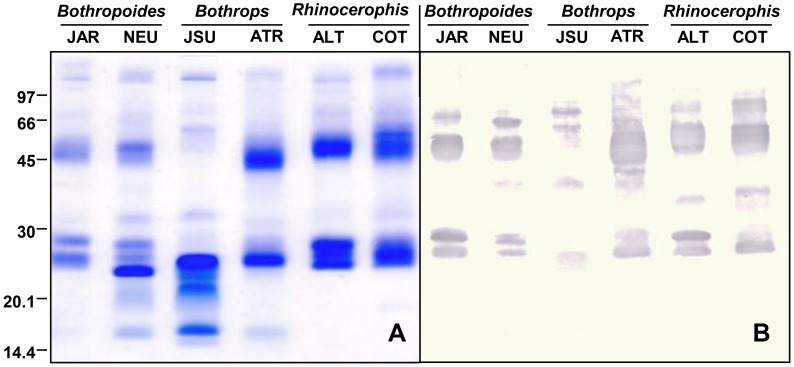
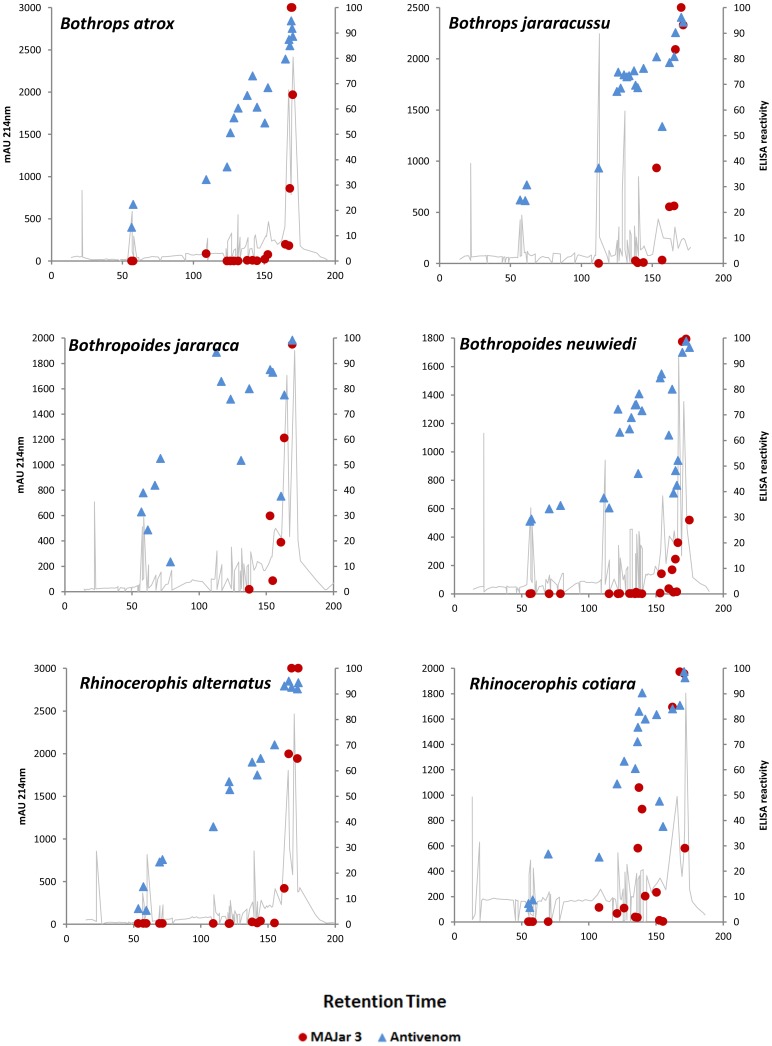
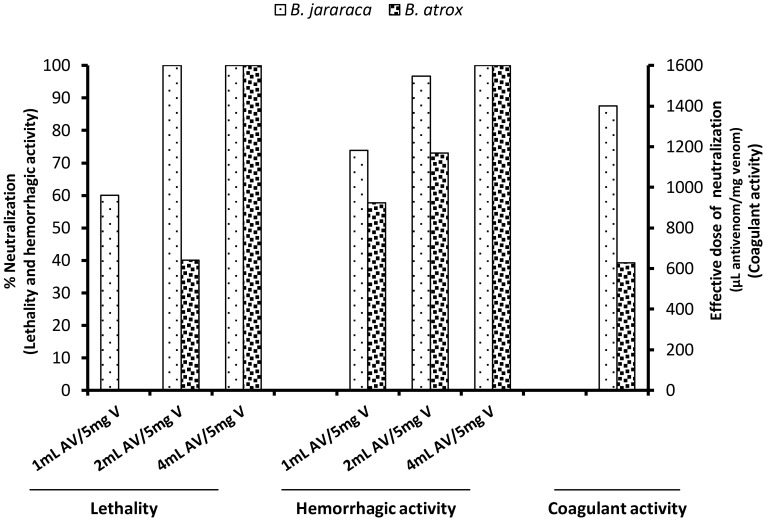
In Latin America, Bothrops snakes account for most snake bites in humans, and the recommended treatment is administration of multispecific Bothrops antivenom (SAB--soro antibotrópico). However, Bothrops snakes are very diverse with regard to their venom composition, which raises the issue of which venoms should be used as immunizing antigens for the production of pan-specific Bothrops antivenoms. In this study, we simultaneously compared the composition and reactivity with SAB of venoms collected from six species of snakes, distributed in pairs from three distinct phylogenetic clades: Bothrops, Bothropoides and Rhinocerophis. We also evaluated the neutralization of Bothrops atrox venom, which is the species responsible for most snake bites in the Amazon region, but not included in the immunization antigen mixture used to produce SAB. Using mass spectrometric and chromatographic approaches, we observed a lack of similarity in protein composition between the venoms from closely related snakes and a high similarity between the venoms of phylogenetically more distant snakes, suggesting little connection between taxonomic position and venom composition. P-III snake venom metalloproteinases (SVMPs) are the most antigenic toxins in the venoms of snakes from the Bothrops complex, whereas class P-I SVMPs, snake venom serine proteinases and phospholipases A2 reacted with antibodies in lower levels. Low molecular size toxins, such as disintegrins and bradykinin-potentiating peptides, were poorly antigenic. Toxins from the same protein family showed antigenic cross-reactivity among venoms from different species; SAB was efficient in neutralizing the B. atrox venom major toxins. Thus, we suggest that it is possible to obtain pan-specific effective antivenoms for Bothrops envenomations through immunization with venoms from only a few species of snakes, if these venoms contain protein classes that are representative of all species to which the antivenom is targeted.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
Pan-American Lancehead Pit-Vipers: Coagulotoxic Venom Effects and Antivenom Neutralisation of Bothrops asper and B. atrox Geographical Variants.Toxins (Basel). 2021 Jan 22;13(2):78. doi: 10.3390/toxins13020078. Toxins (Basel). 2021. PMID: 33499001 Free PMC article.
-
Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management.J Proteomics. 2009 Mar 6;72(2):227-40. doi: 10.1016/j.jprot.2009.01.005. Epub 2009 Jan 16. J Proteomics. 2009. PMID: 19457355
-
Immunological profile of antivenoms: preclinical analysis of the efficacy of a polyspecific antivenom through antivenomics and neutralization assays.J Proteomics. 2014 Jun 13;105:340-50. doi: 10.1016/j.jprot.2014.02.021. Epub 2014 Feb 28. J Proteomics. 2014. PMID: 24583507 Review.
-
Snake venomics and antivenomics of Bothrops atrox venoms from Colombia and the Amazon regions of Brazil, Perú and Ecuador suggest the occurrence of geographic variation of venom phenotype by a trend towards paedomorphism.J Proteomics. 2009 Nov 2;73(1):57-78. doi: 10.1016/j.jprot.2009.07.013. Epub 2009 Aug 6. J Proteomics. 2009. PMID: 19665598
-
Preclinical assessment of the neutralizing efficacy of snake antivenoms in Latin America and the Caribbean: A review.Toxicon. 2018 May;146:138-150. doi: 10.1016/j.toxicon.2018.02.053. Epub 2018 Mar 3. Toxicon. 2018. PMID: 29510161 Review.
Cited by
-
From birth to bite: the evolutionary ecology of India's medically most important snake venoms.BMC Biol. 2024 Jul 29;22(1):161. doi: 10.1186/s12915-024-01960-8. BMC Biol. 2024. PMID: 39075553 Free PMC article.
-
Observation of Bothrops atrox Snake Envenoming Blister Formation from Five Patients: Pathophysiological Insights.Toxins (Basel). 2021 Nov 13;13(11):800. doi: 10.3390/toxins13110800. Toxins (Basel). 2021. PMID: 34822585 Free PMC article.
-
Functional variability of snake venom metalloproteinases: adaptive advantages in targeting different prey and implications for human envenomation.PLoS One. 2014 Oct 14;9(10):e109651. doi: 10.1371/journal.pone.0109651. eCollection 2014. PLoS One. 2014. PMID: 25313513 Free PMC article.
-
Application of an Extracellular Matrix-Mimicking Fluorescent Polymer for the Detection of Proteolytic Venom Toxins.Toxins (Basel). 2023 Apr 18;15(4):294. doi: 10.3390/toxins15040294. Toxins (Basel). 2023. PMID: 37104232 Free PMC article.
-
The Sequence and Three-Dimensional Structure Characterization of Snake Venom Phospholipases B.Front Mol Biosci. 2020 Aug 5;7:175. doi: 10.3389/fmolb.2020.00175. eCollection 2020. Front Mol Biosci. 2020. PMID: 32850964 Free PMC article.
References
-
- WHO (2007) Rabies and envenomings. A neglected public health issue. Available: http://whqlibdoc.who.int/publications/2007/9789241563482_eng.pdf
-
- Ministério-da-Saúde (2001) Manual de diagnóstico e tratamento de acidentes por animais peçonhentos. 2nd ed. Brasília: Fundação Nacional de Saúde.
-
- Brazil V (1918) Do envenenamento ofídico e seu tratamento. Collectanea de Trabalhos (1901–1917) - Instituto Butantan. São Paulo: Typographia do Diário Official. pp. 31–52.
-
- Calvete JJ, Juárez P, Sanz L (2007) Snake venomics. Strategy and applications. J Mass Spectrom 42: 1405–1414. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
