

Characterization, design, and function of the mitochondrial proteome: from organs to organisms
- PMID: 24070373
- PMCID: PMC4076470
- DOI: 10.1021/pr400539j
Characterization, design, and function of the mitochondrial proteome: from organs to organisms
Abstract
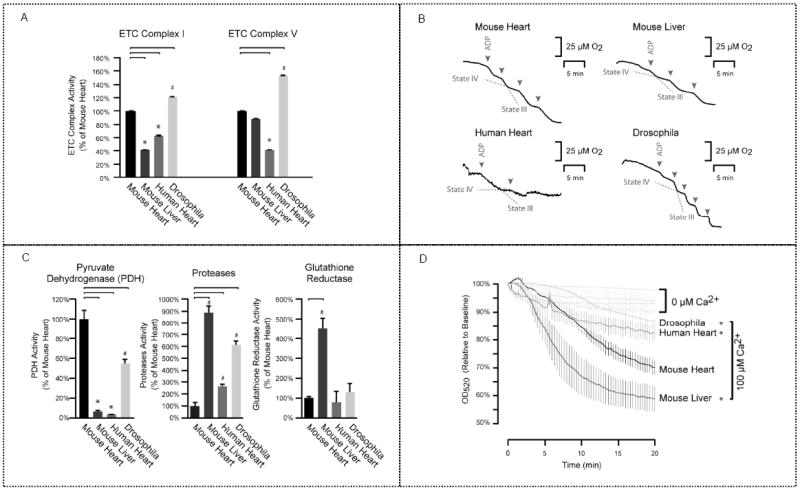
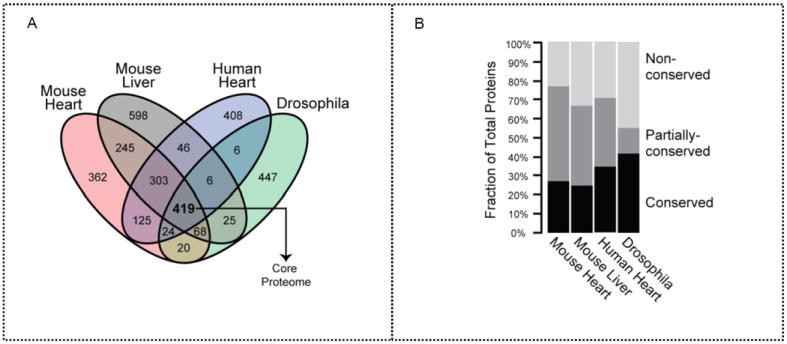
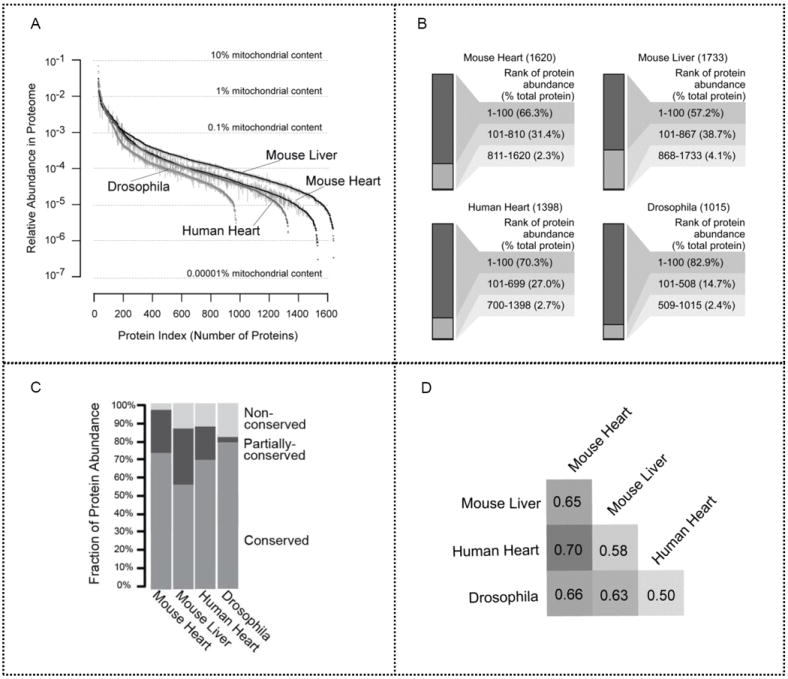
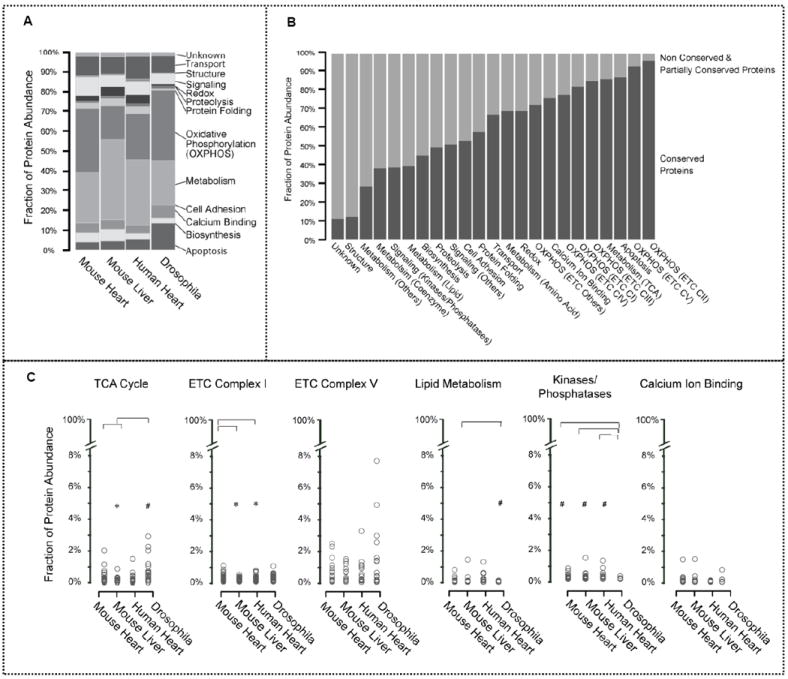
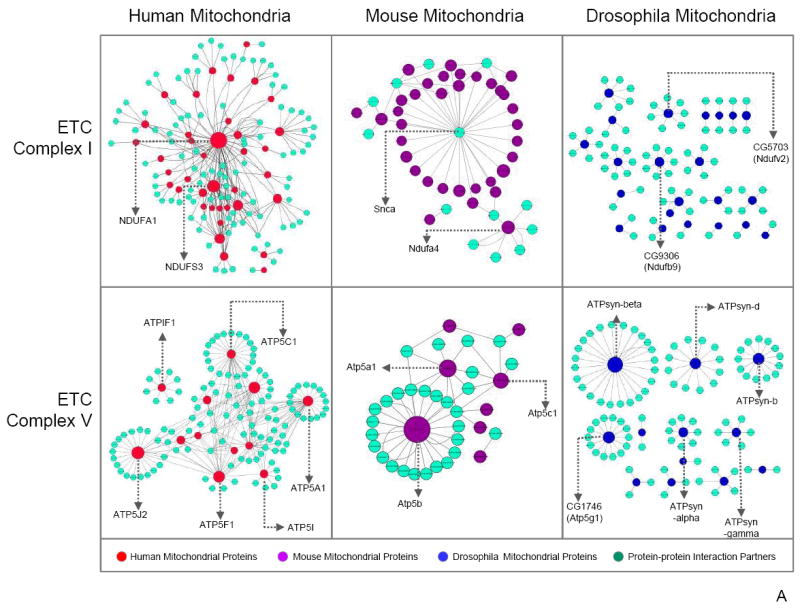
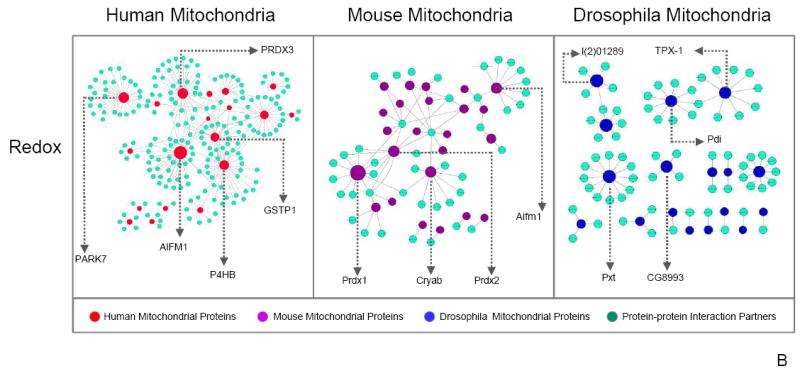
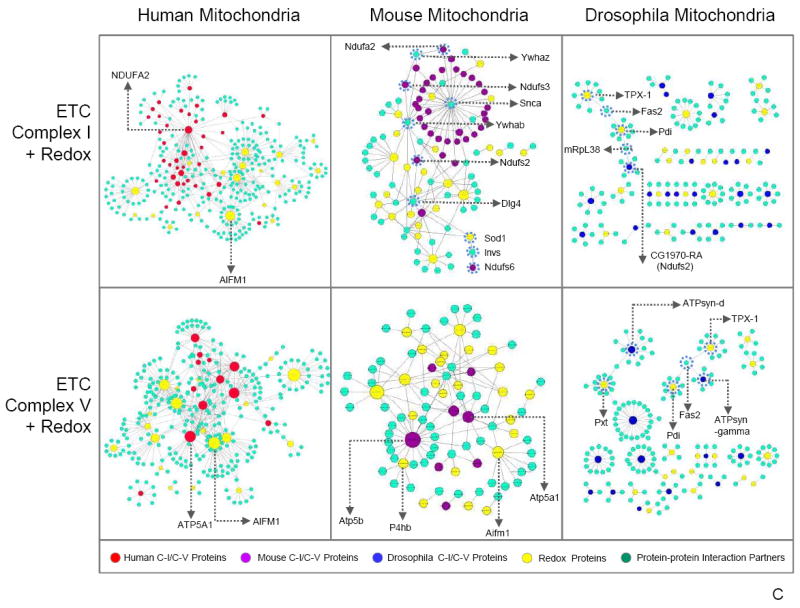
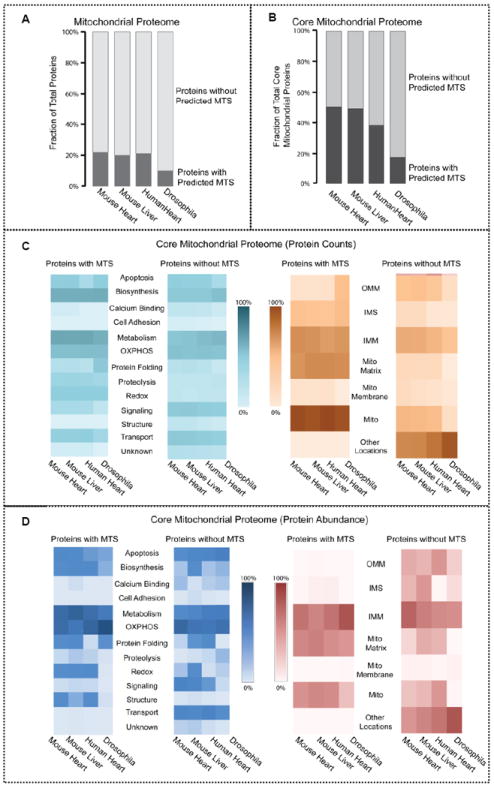
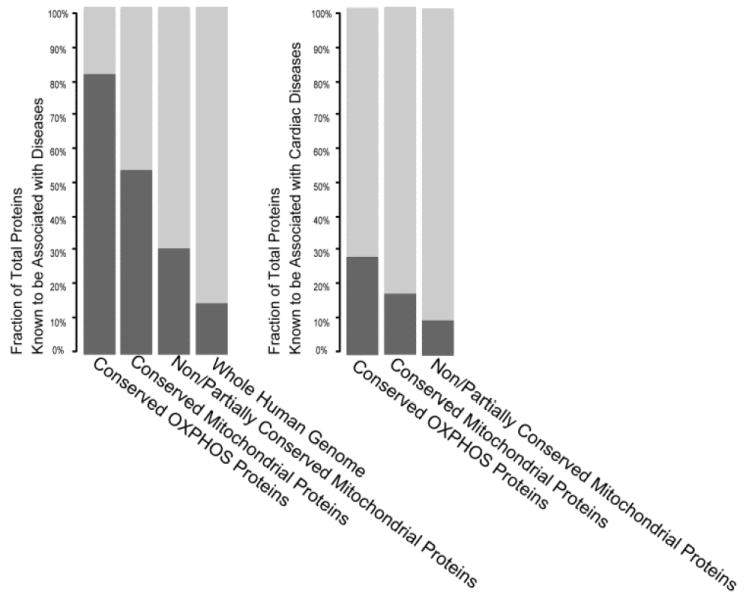
Mitochondria are a common energy source for organs and organisms; their diverse functions are specialized according to the unique phenotypes of their hosting environment. Perturbation of mitochondrial homeostasis accompanies significant pathological phenotypes. However, the connections between mitochondrial proteome properties and function remain to be experimentally established on a systematic level. This uncertainty impedes the contextualization and translation of proteomic data to the molecular derivations of mitochondrial diseases. We present a collection of mitochondrial features and functions from four model systems, including two cardiac mitochondrial proteomes from distinct genomes (human and mouse), two unique organ mitochondrial proteomes from identical genetic codons (mouse heart and mouse liver), as well as a relevant metazoan out-group (drosophila). The data, composed of mitochondrial protein abundance and their biochemical activities, capture the core functionalities of these mitochondria. This investigation allowed us to redefine the core mitochondrial proteome from organs and organisms, as well as the relevant contributions from genetic information and hosting milieu. Our study has identified significant enrichment of disease-associated genes and their products. Furthermore, correlational analyses suggest that mitochondrial proteome design is primarily driven by cellular environment. Taken together, these results connect proteome feature with mitochondrial function, providing a prospective resource for mitochondrial pathophysiology and developing novel therapeutic targets in medicine.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Johnson DT, Harris RA, Blair PV, Balaban RS. Functional consequences of mitochondrial proteome heterogeneity. Am J Physiol : Cell Physiol. 2007;292:C698–707. - PubMed
-
- Balaban RS. Regulation of oxidative phosphorylation in the mammalian cell. Am J Physiol. 1990:377–389. - PubMed
-
- Zhang J, Lin A, Powers J, Lam MP, Lotz C, Liem D, Lau E, Wang D, Deng N, Korge P, Zong NC, Cai H, Weiss J, Ping P. Perspectives on: Sgp symposium on mitochondrial physiology and medicine: Mitochondrial proteome design: From molecular identity to pathophysiological regulation. J Gen Physiol. 2012;139:395–406. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
