

Stabilization of speckle-type POZ protein (Spop) by Daz interacting protein 1 (Dzip1) is essential for Gli turnover and the proper output of Hedgehog signaling
- PMID: 24072710
- PMCID: PMC3820914
- DOI: 10.1074/jbc.M113.512962
Stabilization of speckle-type POZ protein (Spop) by Daz interacting protein 1 (Dzip1) is essential for Gli turnover and the proper output of Hedgehog signaling
Abstract
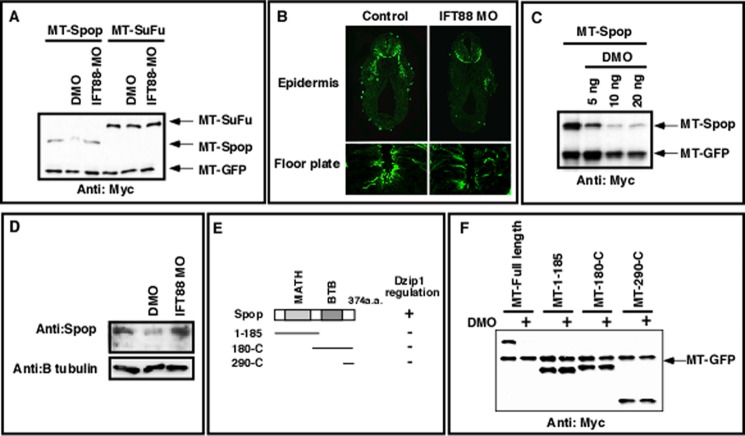
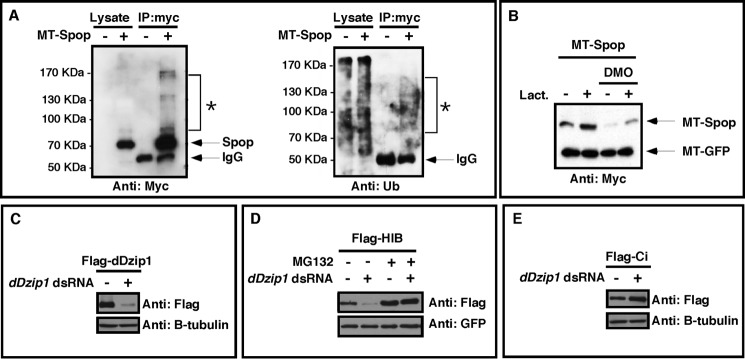
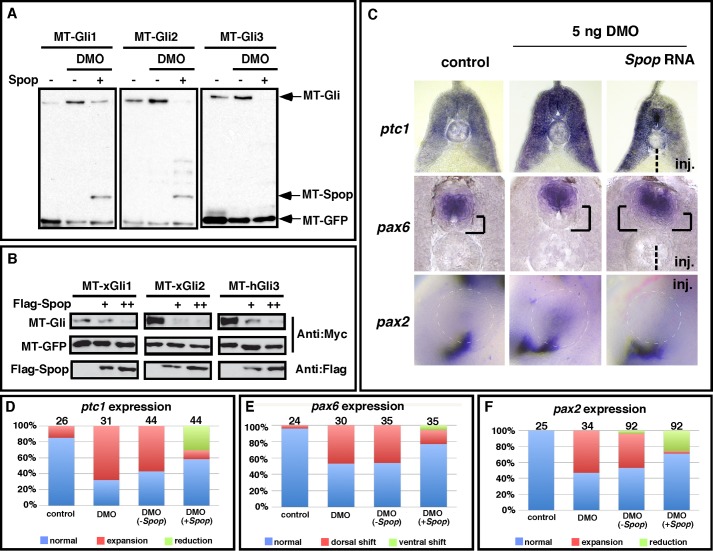
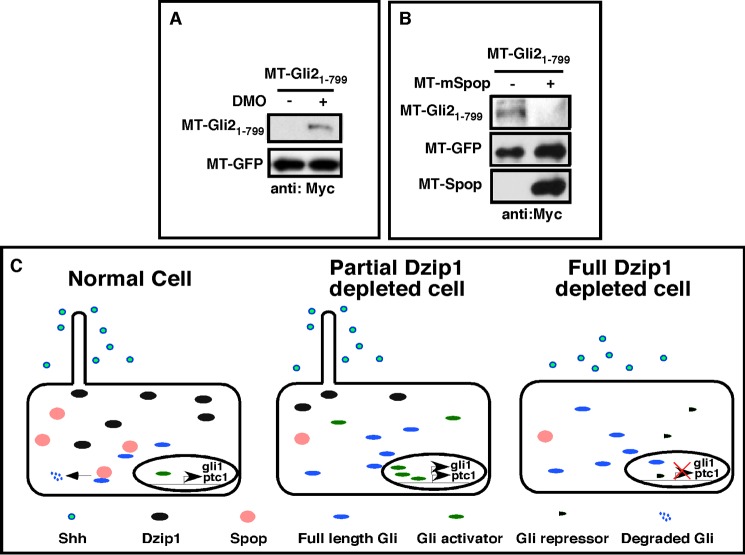
The Hedgehog (Hh) pathway is essential for embryonic development and adult tissue homeostasis. The Gli/Cubitus interruptus (Ci) family of transcription factors acts at the downstream end of the pathway to mediate Hh signaling. Both Hh-dependent and -independent Gli regulatory mechanisms are important for the output of Hh signaling. Daz interacting protein 1 (Dzip1) has bipartite positive and negative functions in the Hh pathway. The positive Hh regulatory function appears to be attributed to a requirement for Dzip1 during ciliogenesis. The mechanism by which Dzip1 inhibits Hh signaling, however, remains largely unclear. We recently found that Dzip1 is required for Gli turnover, which may account for its inhibitory function in Hh signaling. Here, we report that Dzip1 regulates Gli/Ci turnover by preventing degradation of speckle-type POZ protein (Spop), a protein that promotes proteasome-dependent turnover of Gli proteins. We provide evidence that Dzip1 regulates the stability of Spop independent of its function in ciliogenesis. Partial knockdown of Dzip1 to levels insufficient for perturbing ciliogenesis, sensitized Xenopus embryos to Hh signaling, leading to phenotypes that resemble activation of Hh signaling. Importantly, overexpression of Spop was able to restore proper Gli protein turnover and rescue phenotypes in Dzip1-depleted embryos. Consistently, depletion of Dzip1 in Drosophila S2 cells destabilized Hh-induced BTB protein (HIB), the Drosophila homolog of Spop, and increased the level of Ci. Thus, Dzip1-dependent stabilization of Spop/HIB is evolutionarily conserved and essential for proper regulation of Gli/Ci proteins in the Hh pathway.
Keywords: Cilia; Development; Dzip1; Gli; Hedgehog; Protein Degradation; Spop; Xenopus.
Figures
References
-
- Dellovade T., Romer J. T., Curran T., Rubin L. L. (2006) The hedgehog pathway and neurological disorders. Annu. Rev. Neurosci. 29, 539–563 - PubMed
-
- Ingham P. W., McMahon A. P. (2001) Hedgehog signaling in animal development. Paradigms and principles. Genes Dev. 15, 3059–3087 - PubMed
-
- Mullor J. L., Sánchez P., Ruiz i Altaba A. (2002) Pathways and consequences. Hedgehog signaling in human disease. Trends Cell Biol. 12, 562–569 - PubMed
-
- Kasper M., Regl G., Frischauf A. M., Aberger F. (2006) GLI transcription factors. Mediators of oncogenic Hedgehog signalling. Eur. J. Cancer 42, 437–445 - PubMed
-
- Taipale J., Beachy P. A. (2001) The Hedgehog and Wnt signalling pathways in cancer. Nature 411, 349–354 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
