

Characterization of three full-length human nonmuscle myosin II paralogs
- PMID: 24072716
- PMCID: PMC3829186
- DOI: 10.1074/jbc.M113.499848
Characterization of three full-length human nonmuscle myosin II paralogs
Abstract
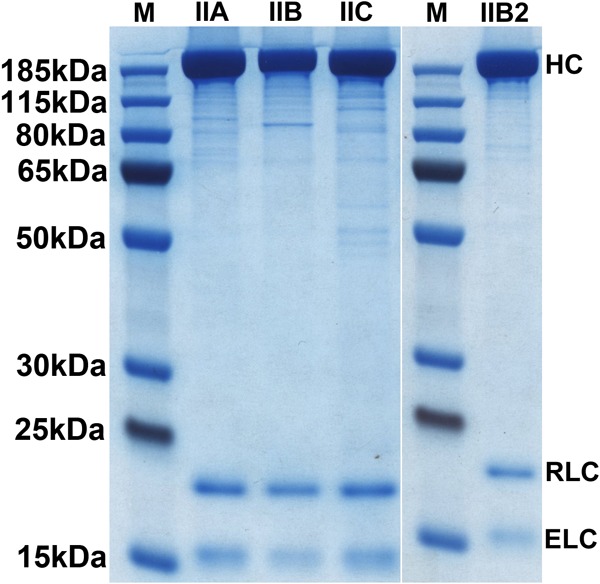
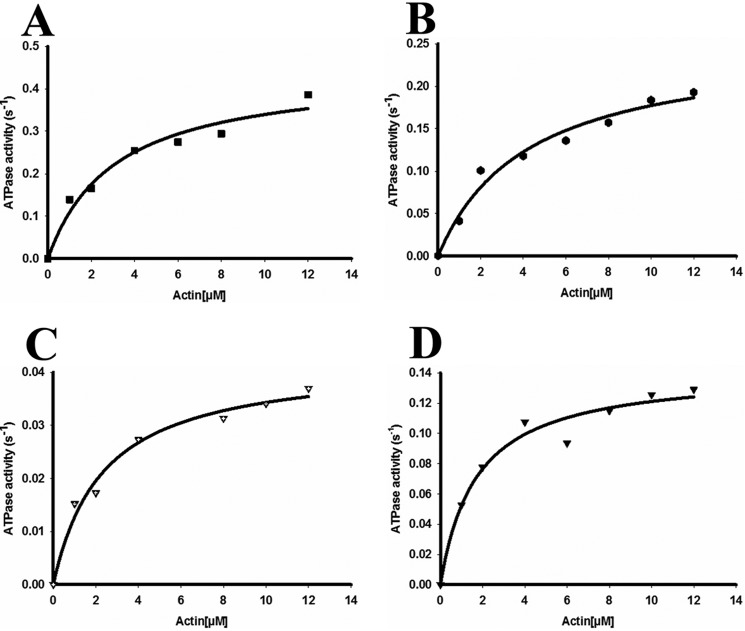
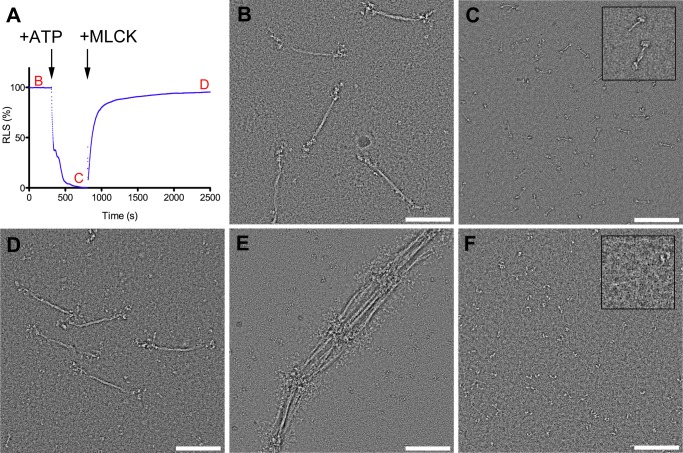
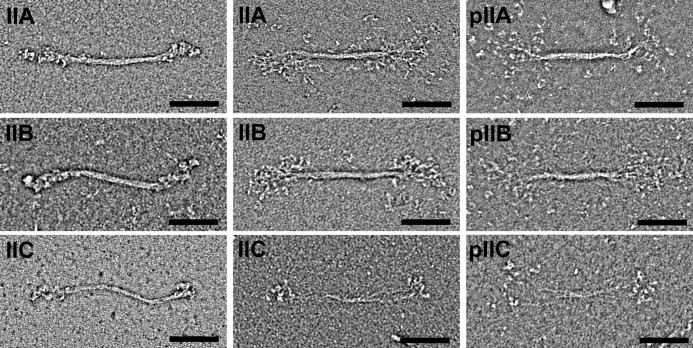
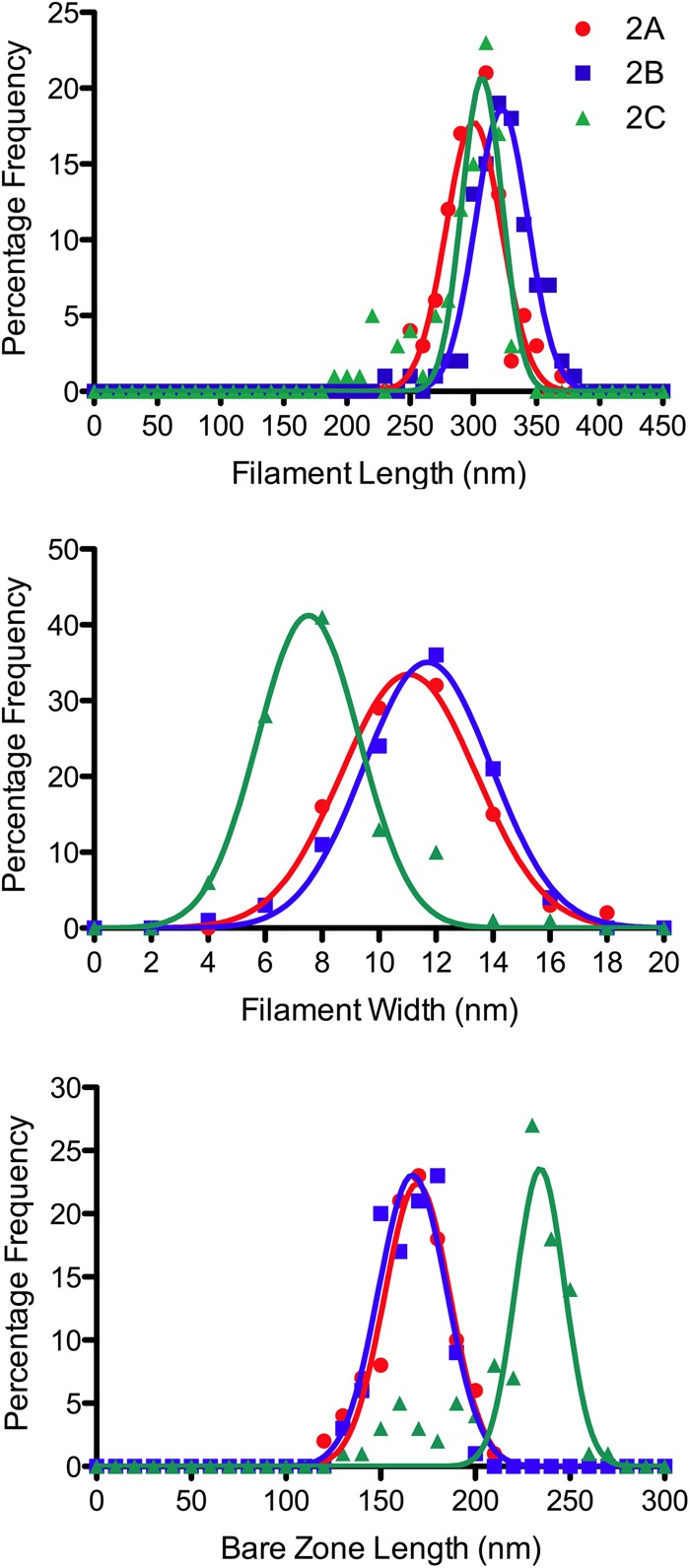
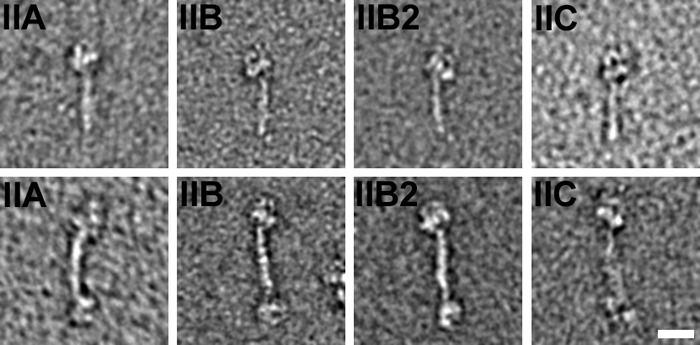
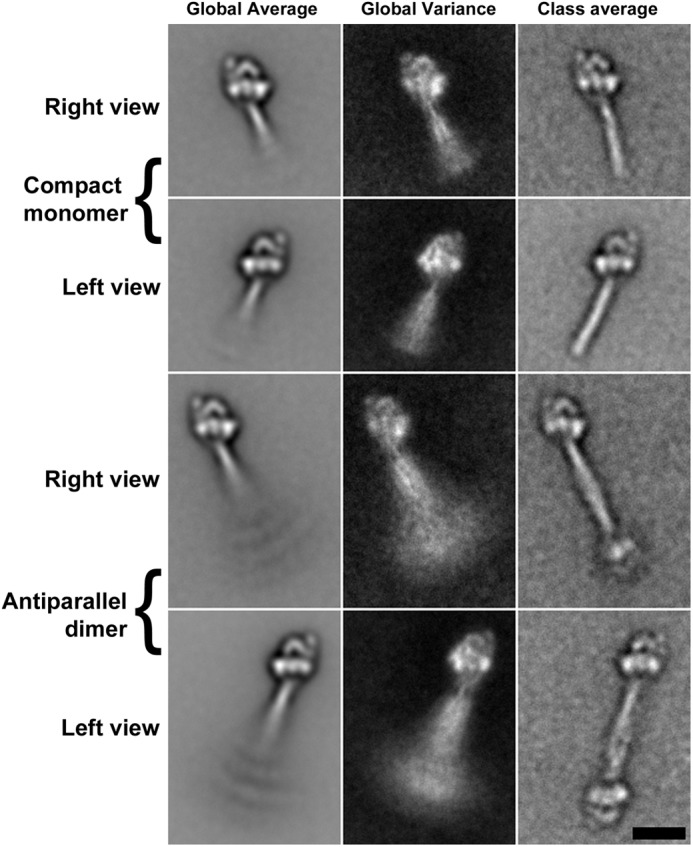
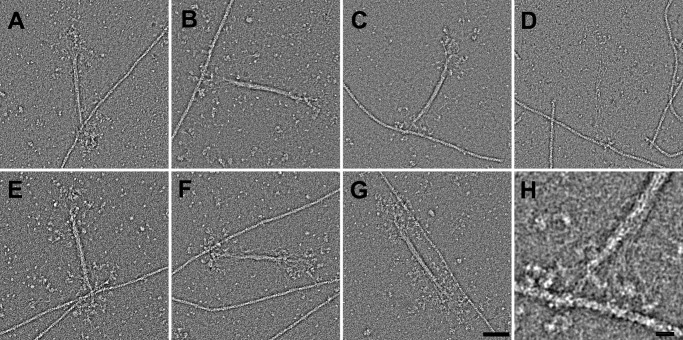
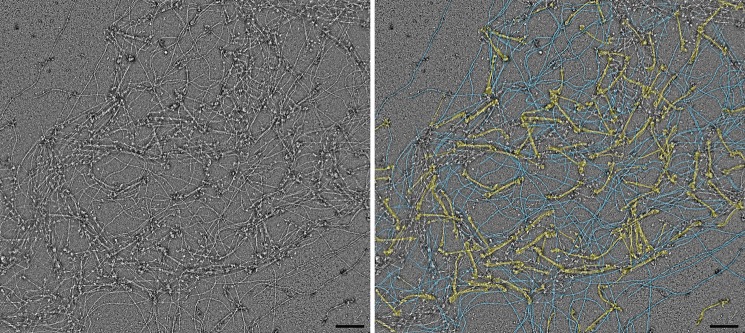
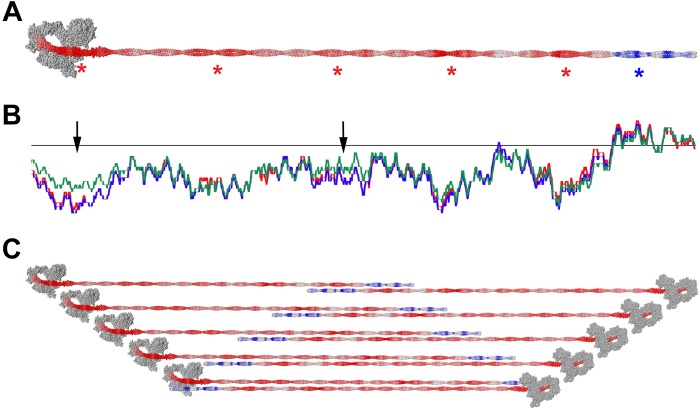
Nonmuscle myosin IIs (NM IIs) are a group of molecular motors involved in a wide variety of cellular processes including cytokinesis, migration, and control of cell morphology. There are three paralogs of the NM II heavy chain in humans (IIA, IIB, and IIC), each encoded by a separate gene. These paralogs are expressed at different levels according to cell type and have different roles and intracellular distributions in vivo. Most previous studies on NM II used tissue-purified protein or expressed fragments of the molecule, which presents potential drawbacks for characterizing individual paralogs of the intact protein in vitro. To circumvent current limitations and approach their native properties, we have successfully expressed and purified the three full-length human NM II proteins with their light chains, using the baculovirus/Sf9 system. The enzymatic and structural properties of the three paralogs were characterized. Although each NM II is capable of forming bipolar filaments, those formed by IIC tend to contain fewer constituent molecules than those of IIA and IIB. All paralogs adopt the compact conformation in the presence of ATP. Phosphorylation of the regulatory light chain leads to assembly into filaments, which bind to actin in the presence of ATP. The nature of interactions with actin filaments is shown with different paralogs exhibiting different actin binding behaviors under equivalent conditions. The data show that although NM IIA and IIB form filaments with similar properties, NM IIC forms filaments that are less well suited to roles such as tension maintenance within the cell.
Keywords: ATPases; Actin; Cytoskeleton; Electron Microscopy (EM); Molecular Motors; Myosin.
Figures
References
-
- Richards T. A., Cavalier-Smith T. (2005) Myosin domain evolution and the primary divergence of eukaryotes. Nature 436, 1113–1118 - PubMed
-
- Kühne W. (1864) Untersuchungen über das protplasma und di contractilitat, W. Engelmann, Leipzig, Germany
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
