

Inhibition of protein translation as a novel mechanism for prostaglandin E2 regulation of cell functions
- PMID: 24072780
- PMCID: PMC3868831
- DOI: 10.1096/fj.13-231720
Inhibition of protein translation as a novel mechanism for prostaglandin E2 regulation of cell functions
Abstract
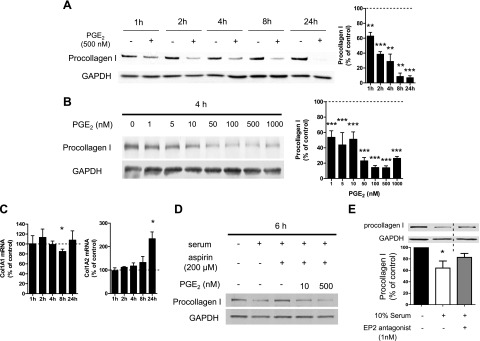
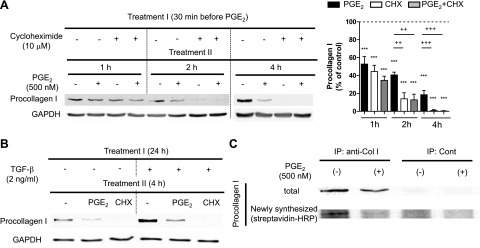
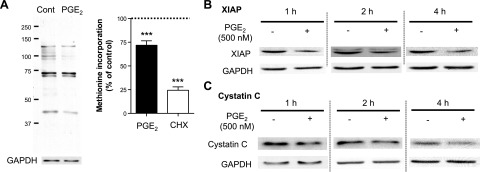
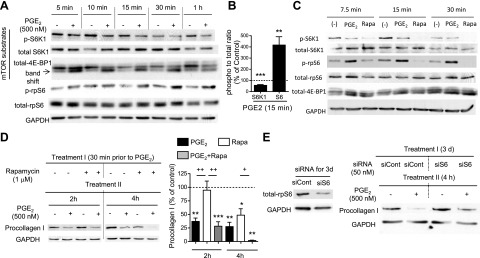
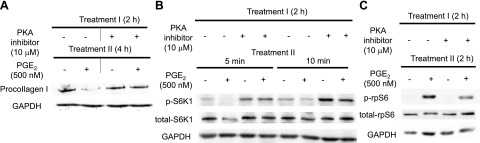
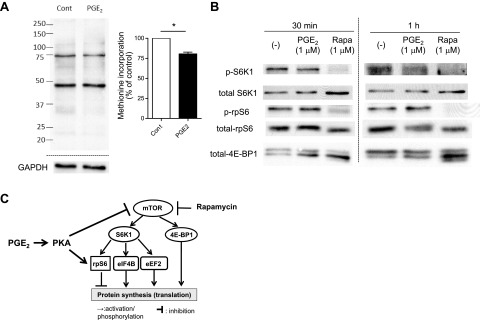
Prostaglandin E2 (PGE2) regulates numerous biological processes by modulating transcriptional activation, epigenetic control, proteolysis, and secretion of various proteins. Scar formation depends on fibroblast elaboration of matrix proteins such as collagen, and this process is strongly suppressed by PGE2 through activation of cAMP-dependent protein kinase A (PKA). However, the actual mechanism by which PGE2-PKA signaling inhibits collagen expression in fibroblasts has never been delineated, and that was the objective of this study. PGE2 unexpectedly induced a rapid reduction in procollagen I protein expression in adult lung fibroblasts, with a half-maximum effect at 1.5 h. This effect reflected its inhibition of translation rather than transcription. Global protein synthesis was also inhibited by PGE2. This action was mediated by PKA and involved both activation of ribosomal protein (rpS6) and suppression of mammalian target of rapamycin (mTOR). Similar effects of PGE2 were demonstrated in mouse peritoneal macrophages (PMs). These findings identify inhibition of translation as a new mechanism by which PGE2 regulates cellular function and a novel example of translational inhibition mediated by opposing actions on two distinct translational control pathways. Translational inhibition would be expected to contribute to dynamic alterations in cell function that accompany the changing PGE2 levels observed in disease states and with various pharmacotherapies.
Keywords: fibroblasts; macrophages; mammalian target of rapamycin; protein kinase A; ribosomal protein S6.
Figures
References
-
- Hetzel M., Bachem M., Anders D., Trischler G., Faehling M. (2005) Different effects of growth factors on proliferation and matrix production of normal and fibrotic human lung fibroblasts. Lung 183, 225–237 - PubMed
-
- Elias J. A., Rossman M. D., Zurier R. B., Daniele R. P. (1985) Human alveolar macrophage inhibition of lung fibroblast growth: a prostaglandin-dependent process. Am. Rev. Respir. Dis. 131, 94–99 - PubMed
-
- Fine A., Poliks C. F., Donahue L. P., Smith B. D., Goldstein R. H. (1989) The differential effect of prostaglandin E2 on transforming growth factor-beta and insulin-induced collagen formation in lung fibroblasts. J. Biol. Chem. 264, 16988–16991 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
