

3' UTR-isoform choice has limited influence on the stability and translational efficiency of most mRNAs in mouse fibroblasts
- PMID: 24072873
- PMCID: PMC3847777
- DOI: 10.1101/gr.156919.113
3' UTR-isoform choice has limited influence on the stability and translational efficiency of most mRNAs in mouse fibroblasts
Abstract
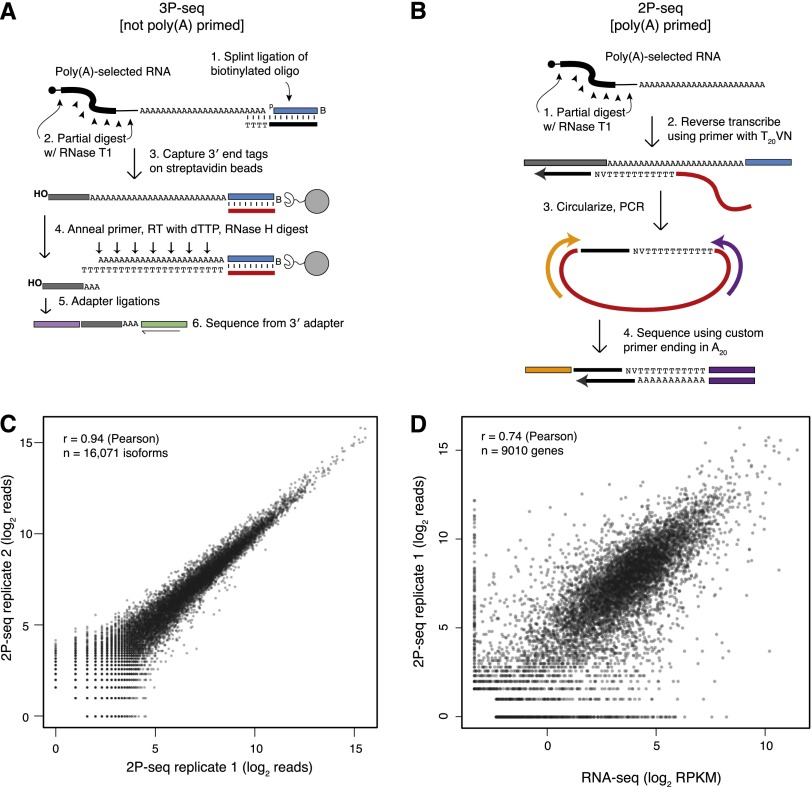
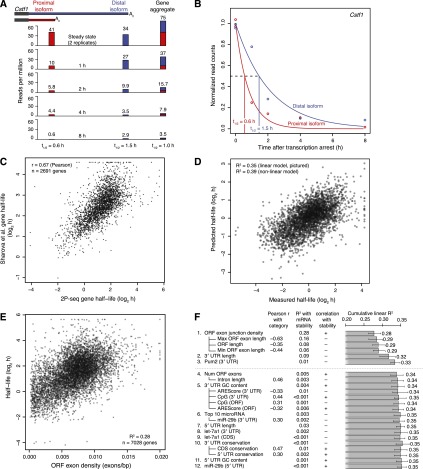
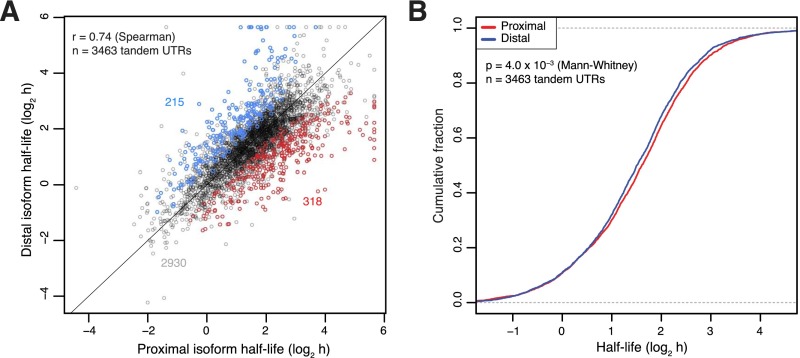
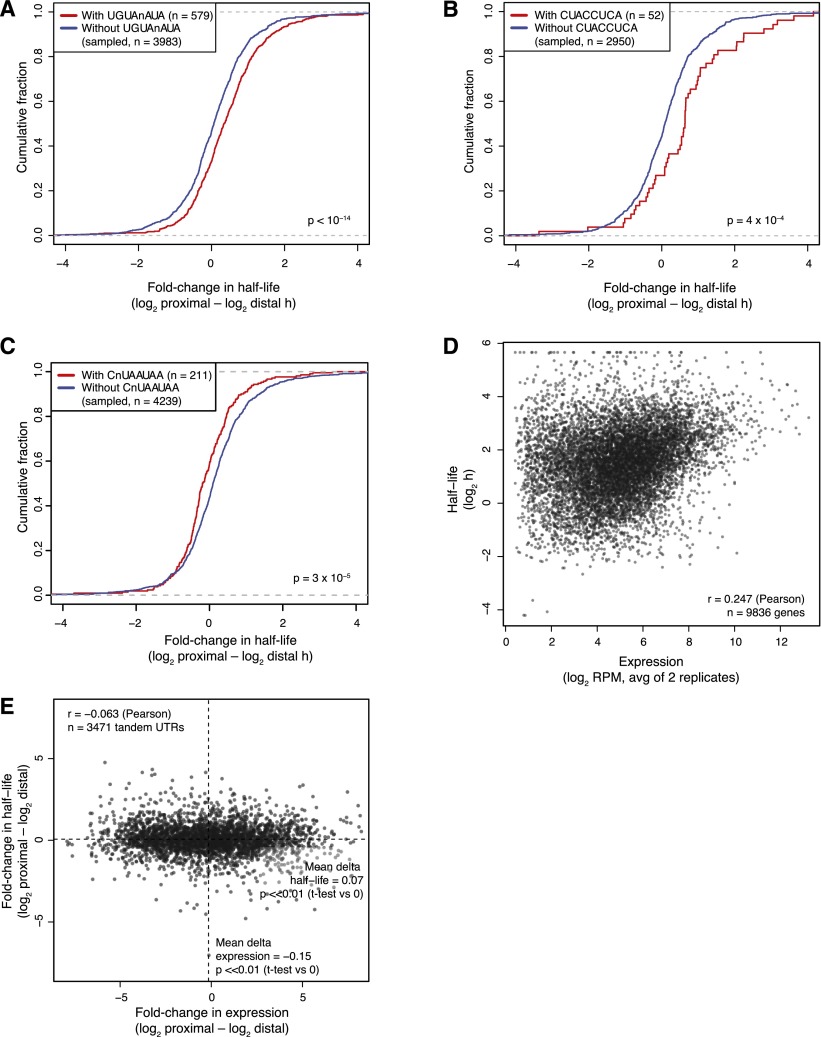
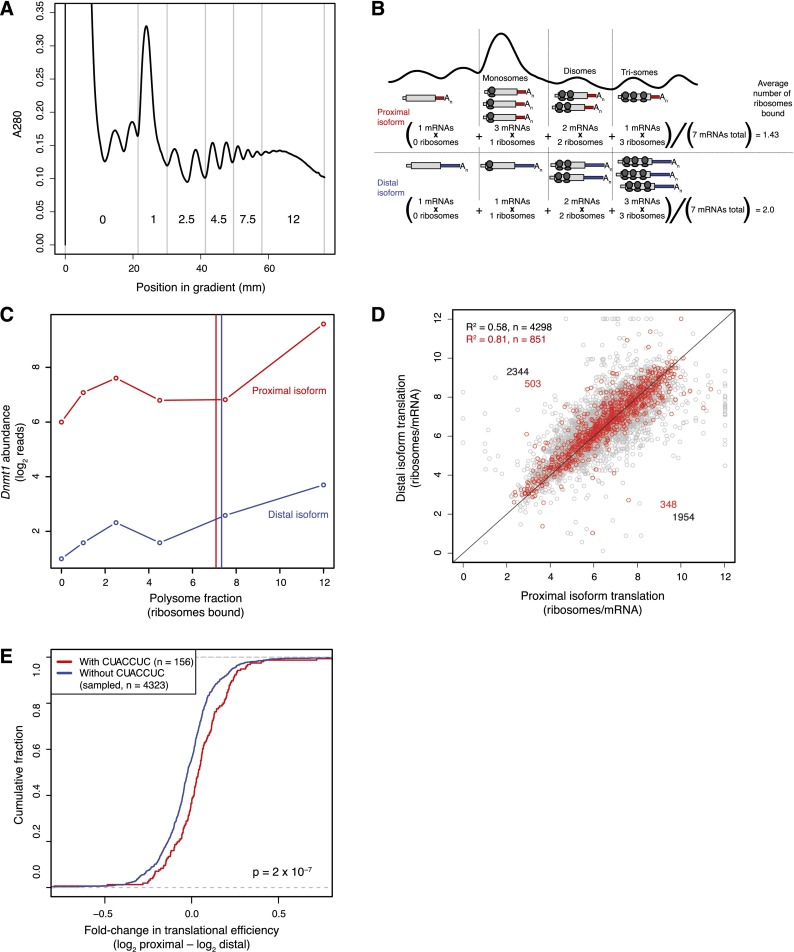
Variation in protein output across the genome is controlled at several levels, but the relative contributions of different regulatory mechanisms remain poorly understood. Here, we obtained global measurements of decay and translation rates for mRNAs with alternative 3' untranslated regions (3' UTRs) in murine 3T3 cells. Distal tandem isoforms had slightly but significantly lower mRNA stability and greater translational efficiency than proximal isoforms on average. The diversity of alternative 3' UTRs also enabled inference and evaluation of both positively and negatively acting cis-regulatory elements. The 3' UTR elements with the greatest implied influence were microRNA complementary sites, which were associated with repression of 32% and 4% at the stability and translational levels, respectively. Nonetheless, both the decay and translation rates were highly correlated for proximal and distal 3' UTR isoforms from the same genes, implying that in 3T3 cells, alternative 3' UTR sequences play a surprisingly small regulatory role compared to other mRNA regions.
Figures
References
-
- Andreassi C, Riccio A 2009. To localize or not to localize: mRNA fate is in 3′UTR ends. Trends Cell Biol 19: 465–474 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases