

Indoxacarb, Metaflumizone, and Other Sodium Channel Inhibitor Insecticides: Mechanism and Site of Action on Mammalian Voltage-Gated Sodium Channels
- PMID: 24072940
- PMCID: PMC3780446
- DOI: 10.1016/j.pestbp.2013.03.004
Indoxacarb, Metaflumizone, and Other Sodium Channel Inhibitor Insecticides: Mechanism and Site of Action on Mammalian Voltage-Gated Sodium Channels
Abstract
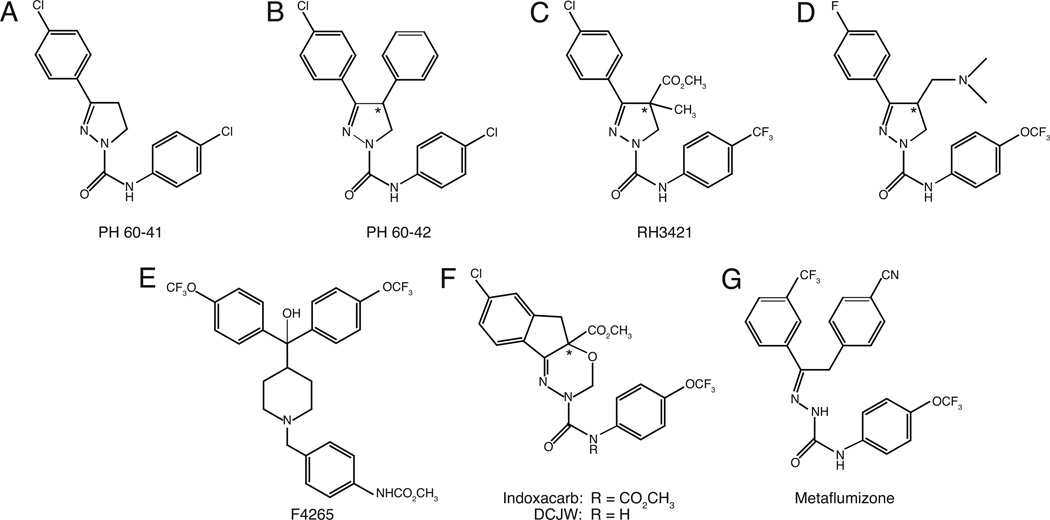
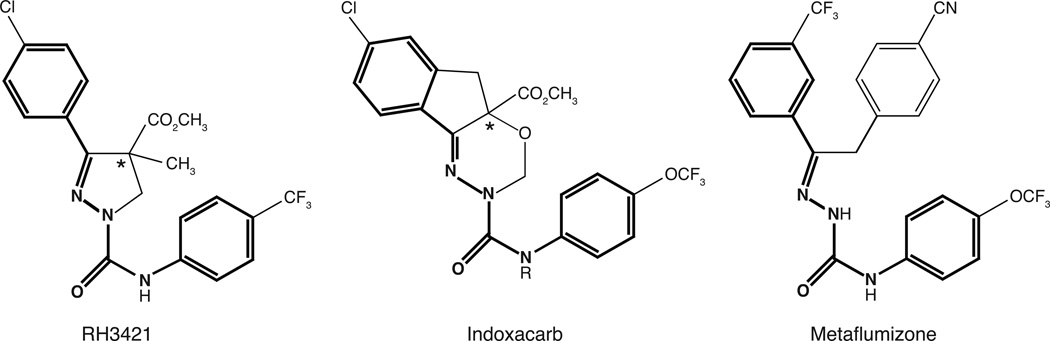
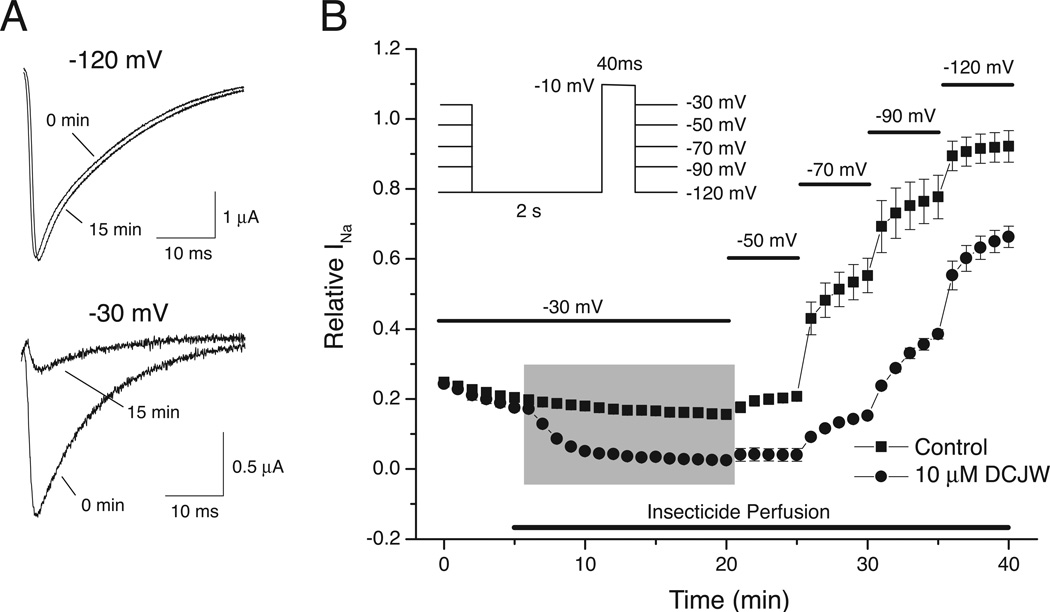
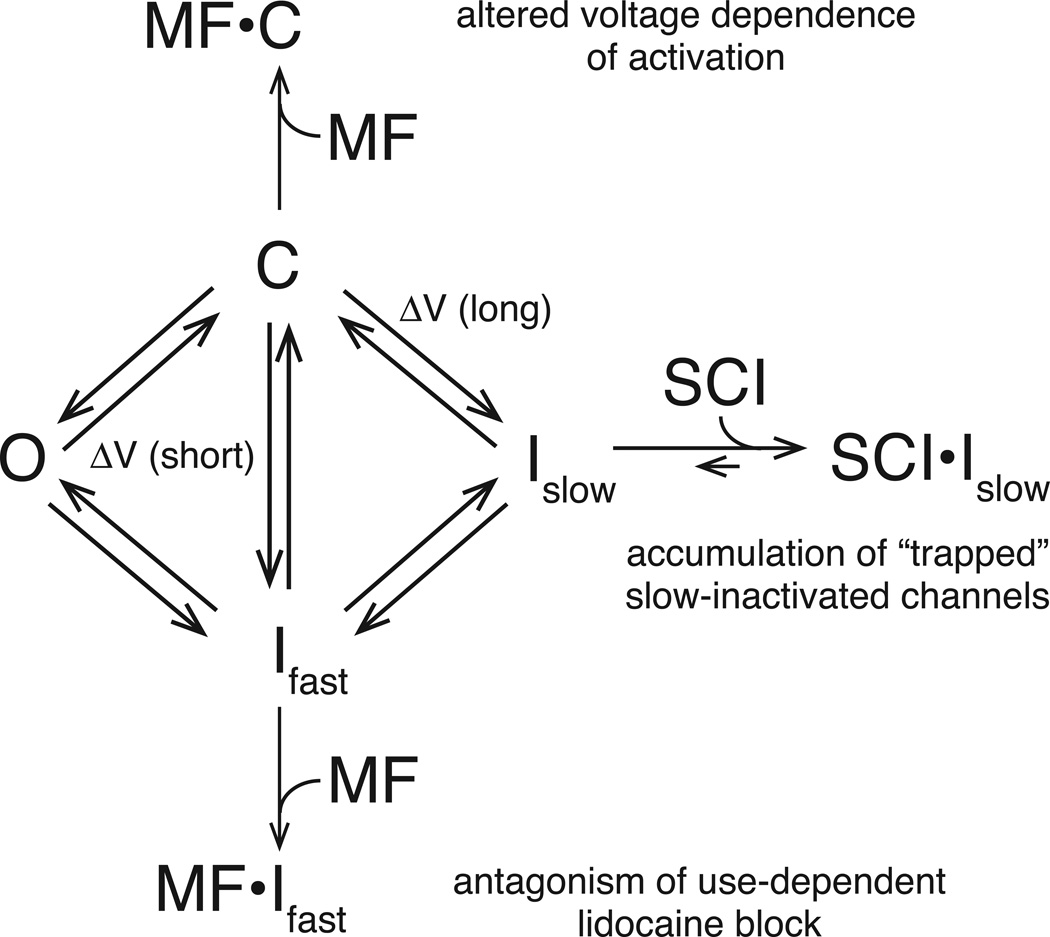
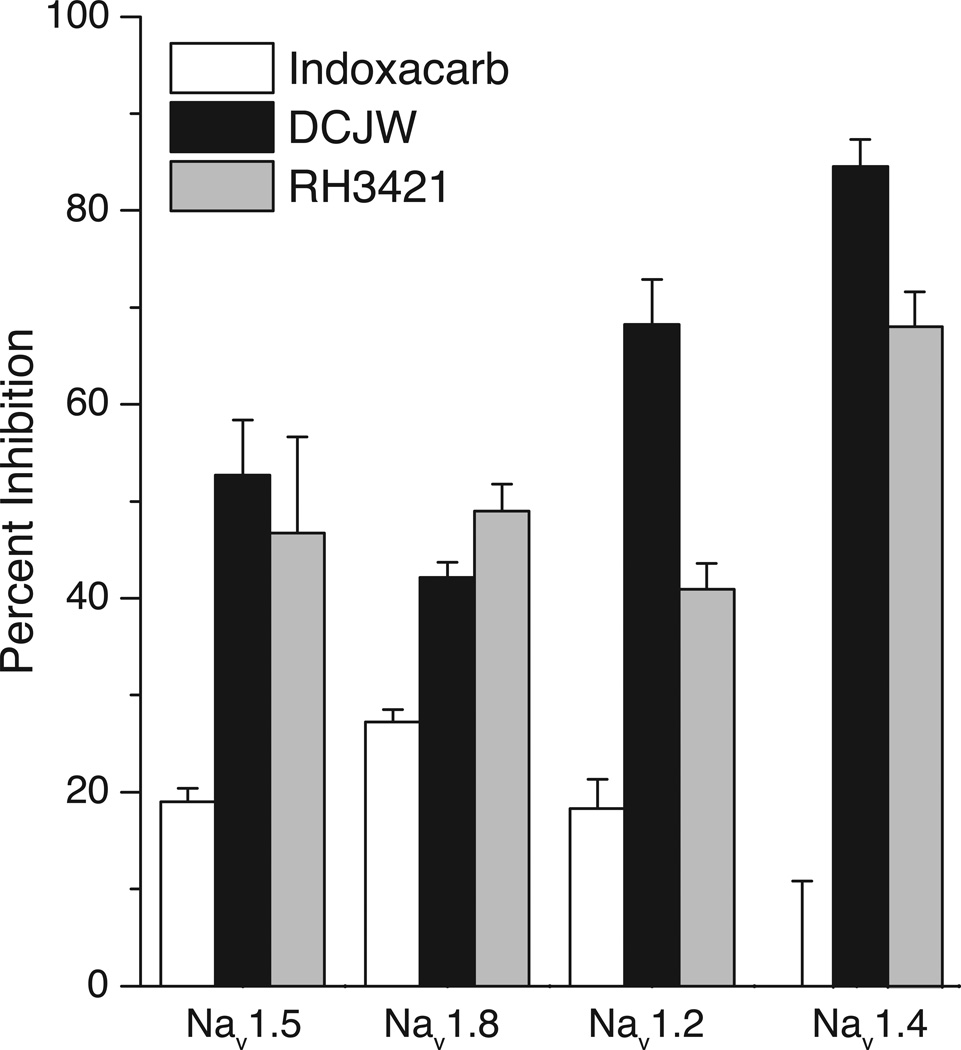
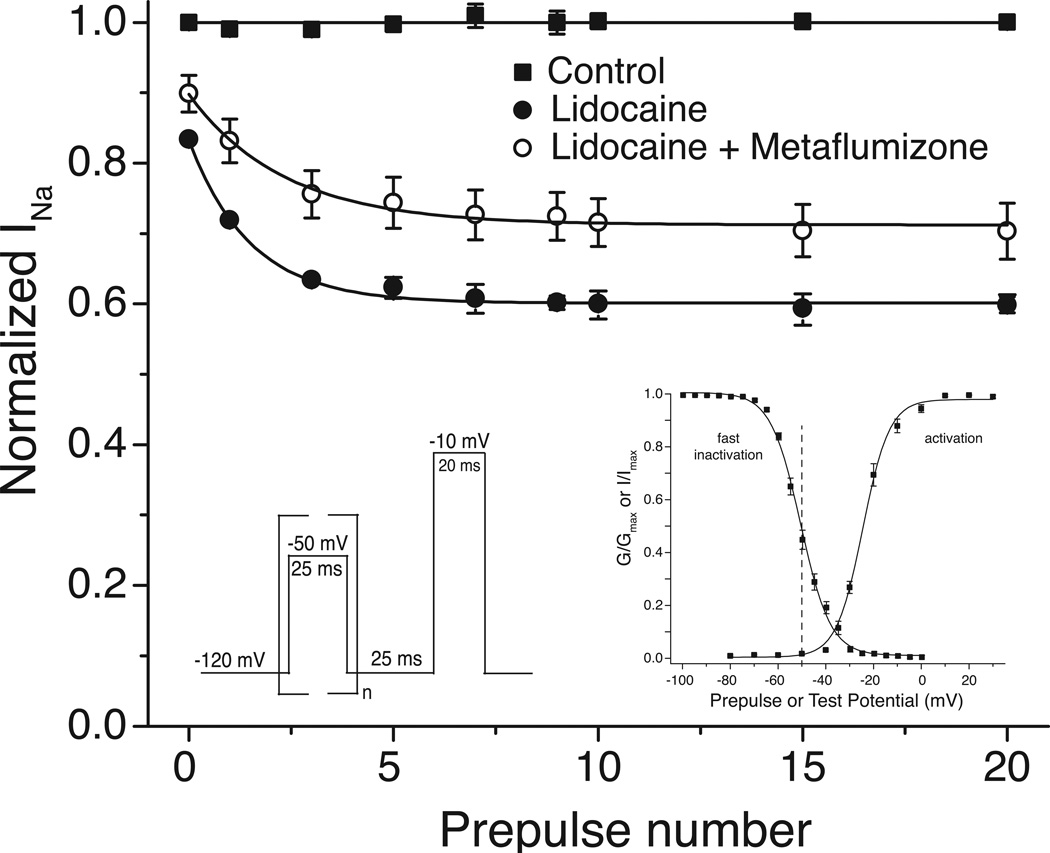
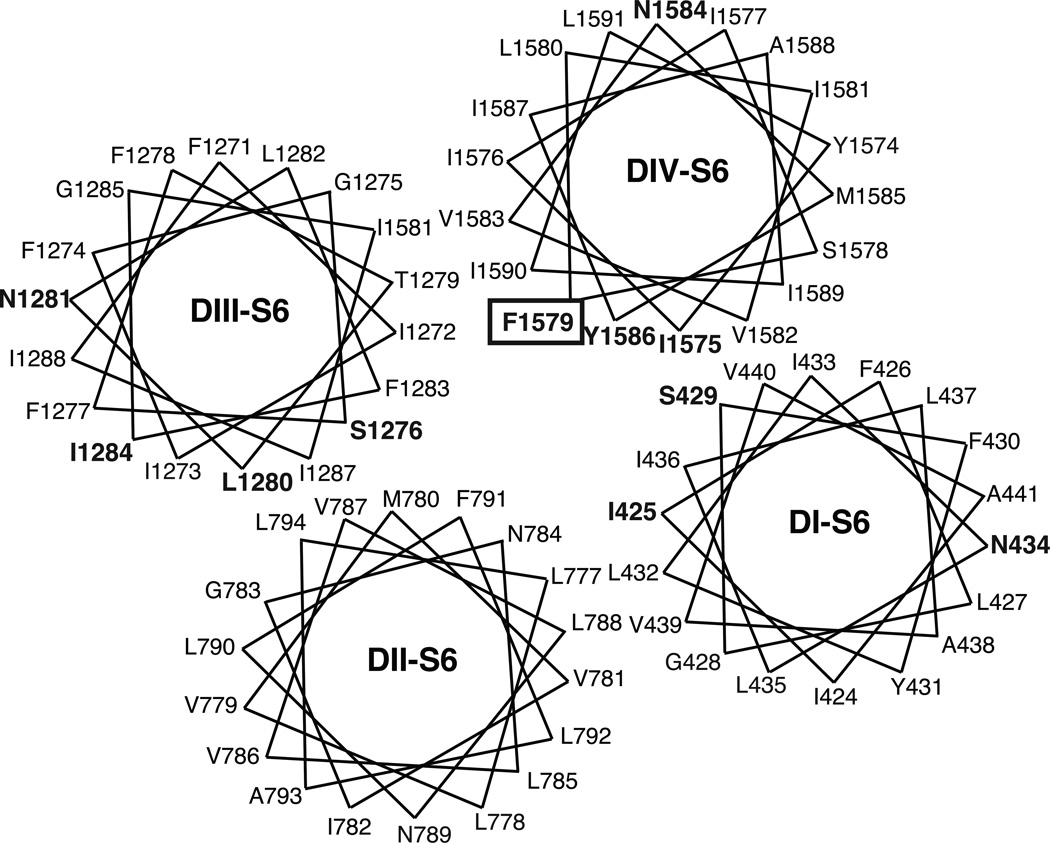
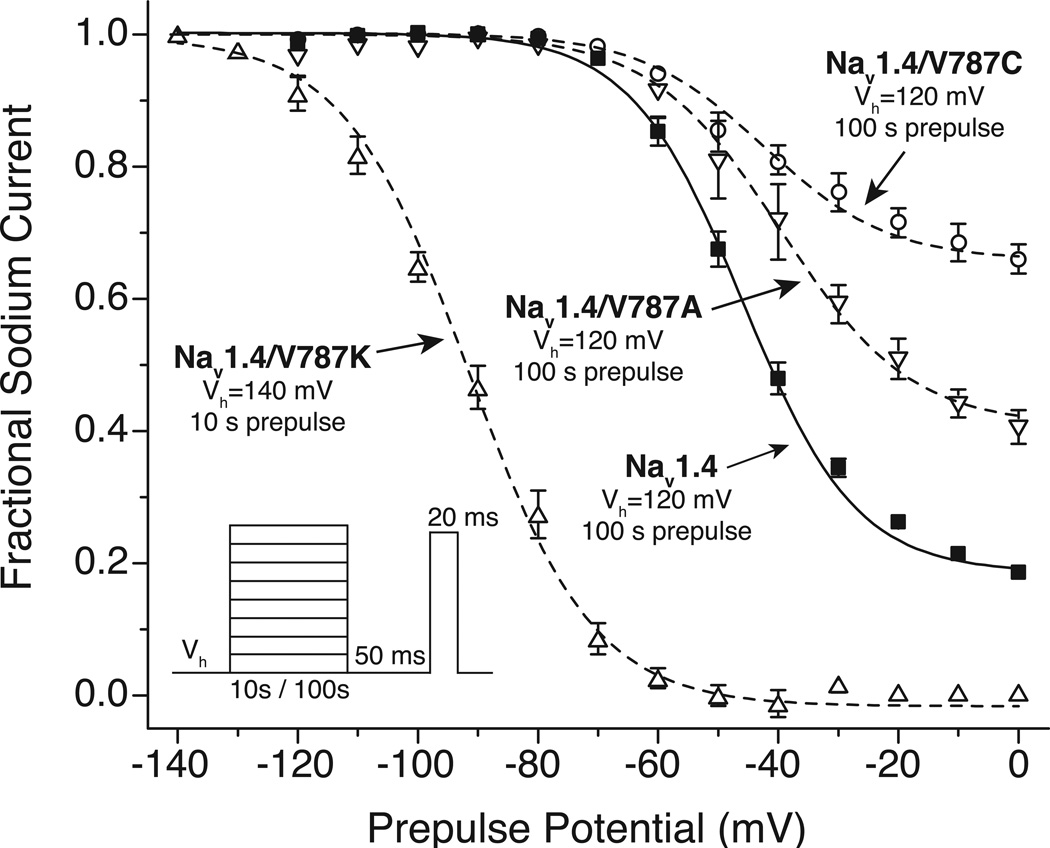
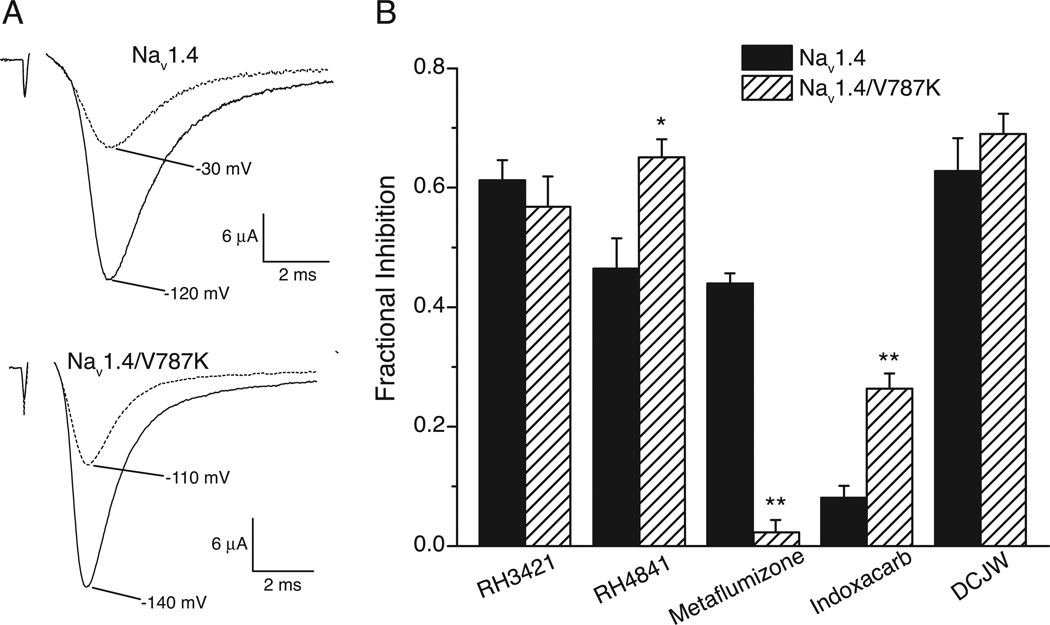
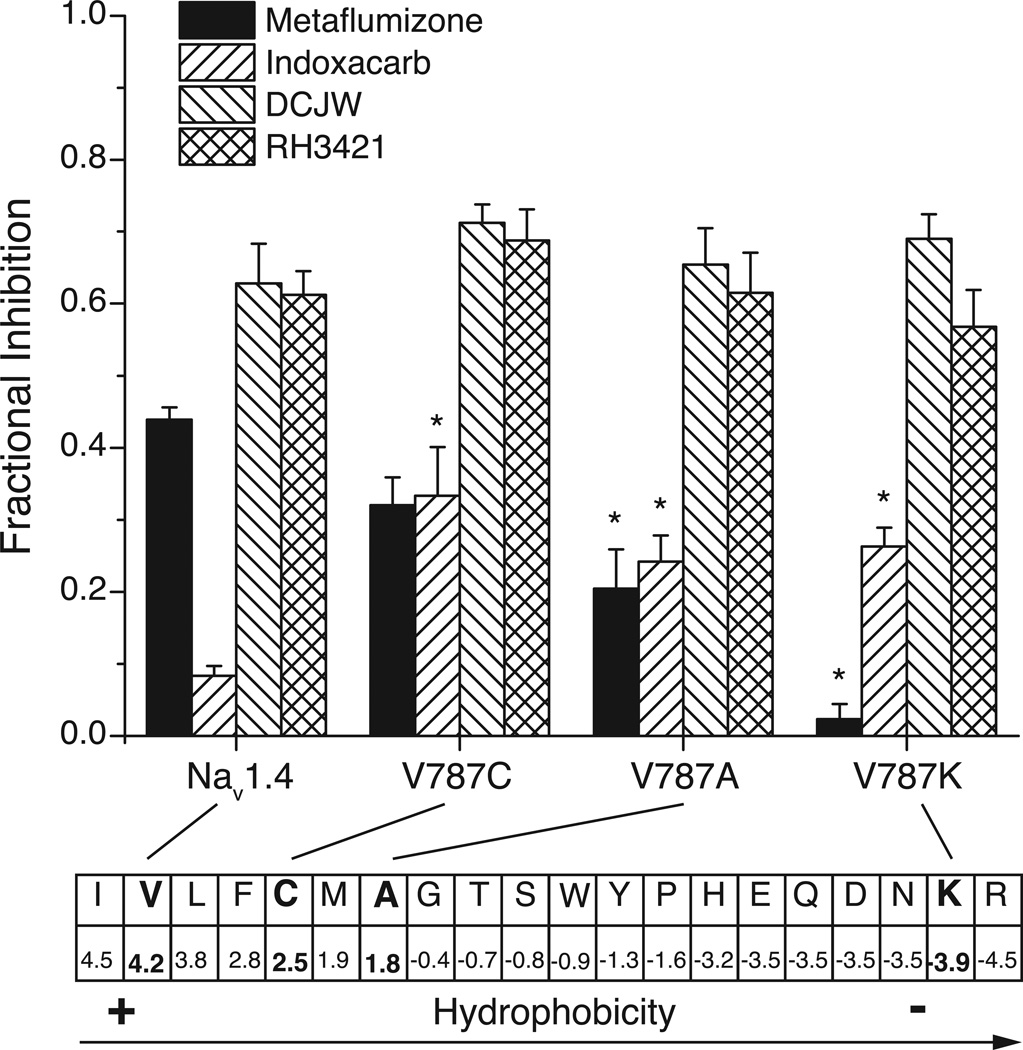
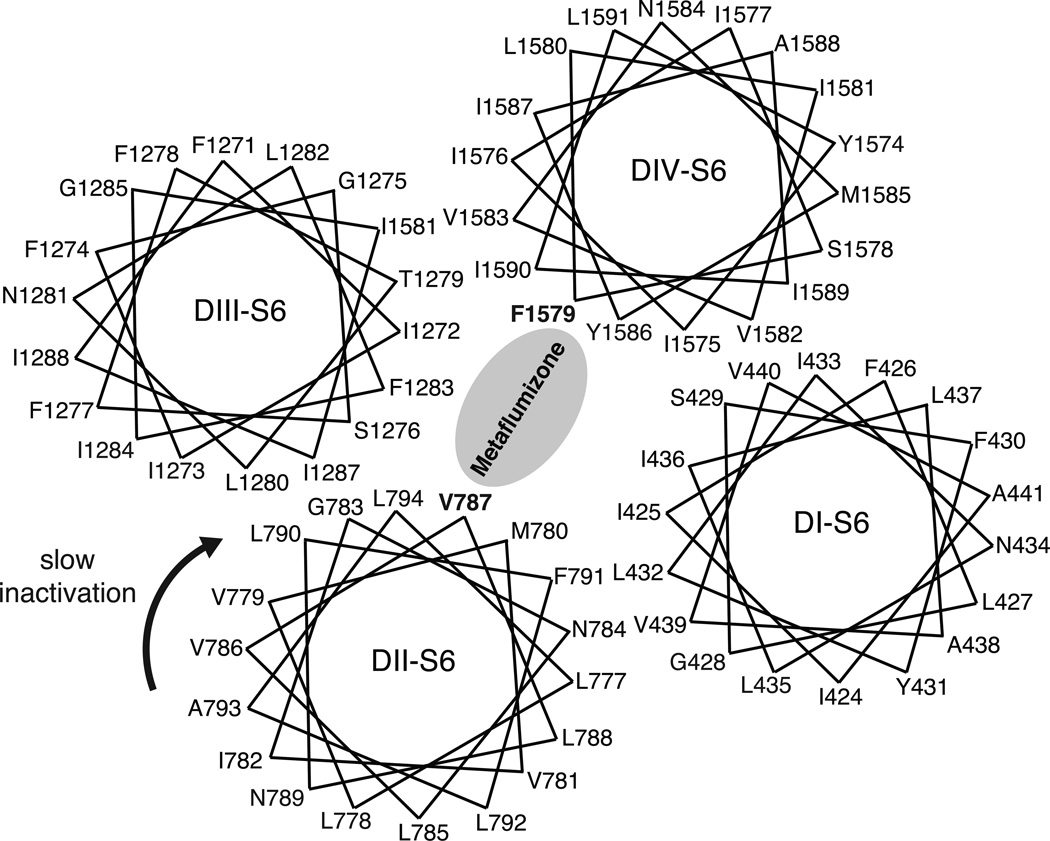
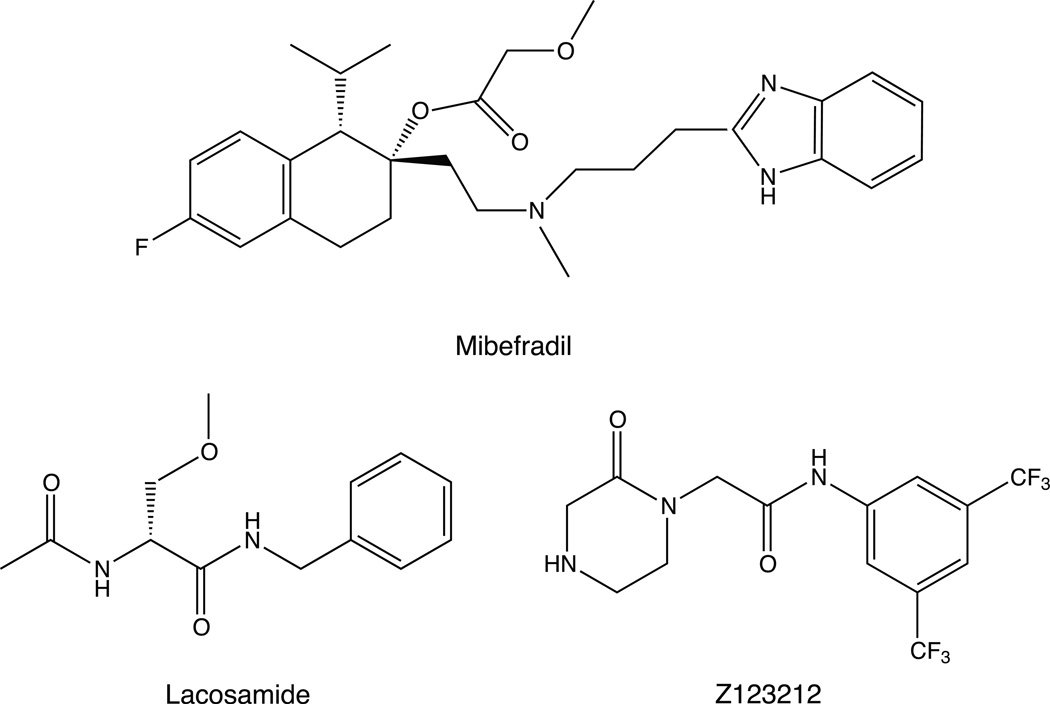
Sodium channel inhibitor (SCI) insecticides were discovered almost four decades ago but have only recently yielded important commercial products (eg., indoxacarb and metaflumizone). SCI insecticides inhibit sodium channel function by binding selectively to slow-inactivated (non-conducting) sodium channel states. Characterization of the action of SCI insecticides on mammalian sodium channels using both biochemical and electrophysiological approaches demonstrates that they bind at or near a drug receptor site, the "local anesthetic (LA) receptor." This mechanism and site of action on sodium channels differentiates SCI insecticides from other insecticidal agents that act on sodium channels. However, SCI insecticides share a common mode of action with drugs currently under investigation as anticonvulsants and treatments for neuropathic pain. In this paper we summarize the development of the SCI insecticide class and the evidence that this structurally diverse group of compounds have a common mode of action on sodium channels. We then review research that has used site-directed mutagenesis and heterologous expression of cloned mammalian sodium channels in Xenopus laevis oocytes to further elucidate the site and mechanism of action of SCI insecticides. The results of these studies provide new insight into the mechanism of action of SCI insecticides on voltage-gated sodium channels, the location of the SCI insecticide receptor, and its relationship to the LA receptor that binds therapeutic SCI agents.
Keywords: DCJW; RH3421; indoxacarb; local anesthetic receptor; metaflumizone; sodium channel.
Figures
References
-
- Mulder R, Wellinga K, van Daalen JJ. A new class of insecticides. Naturwissenschaften. 1975;62:531–532. - PubMed
-
- McCann SF, Annis GD, Shapiro R, Piotrowski DW, Lahm GP, Long JK, Lee KC, Hughes MM, Myers BJ, Griswold SM, Reeves BW, March RW, Sharpe PL, Lowder P, Barnette WE, Wing KD. The discovery of indoxacarb: oxadiazines as a new class of pyrazoline-type insecticide. Pest Management Science. 2001;57:153–164. - PubMed
-
- Meier GA, Silverman R, Ray PS, Cullen TG, Ali SF, Marek FL, Webster CA. Insecticidal dihydropyrazoles with reduced lipophilicity. In: Baker DR, Fenyes JG, Steffens JJ, editors. Synthesis and Chemistry of Agrochemicals III. Washington, DC: American Chemical Society; 1992. pp. 313–326.
-
- Takagi K, Hamaguchi H, Nishimatsu T, Konno T. Discovery of metaflumizonea novel semicarbazone insecticide. Veterinary Parasitology. 2007;150:177–181. - PubMed
-
- Silver KS, Soderlund DM. Action of pyrazoline-type insecticides at neuronal target sites. Pesticide Biochemistry and Physiology. 2005;81:136–143.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources