

MiRNAs confer phenotypic robustness to gene networks by suppressing biological noise
- PMID: 24077216
- PMCID: PMC3836244
- DOI: 10.1038/ncomms3364
MiRNAs confer phenotypic robustness to gene networks by suppressing biological noise
Abstract
miRNAs are small non-coding RNAs able to modulate target gene expression. It has been postulated that miRNAs confer robustness to biological processes, but clear experimental evidence is still missing. Here, using a synthetic biological approach, we demonstrate that microRNAs provide phenotypic robustness to transcriptional regulatory networks by buffering fluctuations in protein levels. We construct a network motif in mammalian cells exhibiting a 'toggle-switch' phenotype in which two alternative protein expression levels define its ON and OFF states. The motif consists of an inducible transcription factor that self-regulates its own transcription and that of a miRNA against the transcription factor itself. We confirm, using mathematical modelling and experimental approaches, that the microRNA confers robustness to the toggle-switch by enabling the cell to maintain and transmit its state. When absent, a dramatic increase in protein noise level occurs, causing the cell to randomly switch between the two states.
Figures
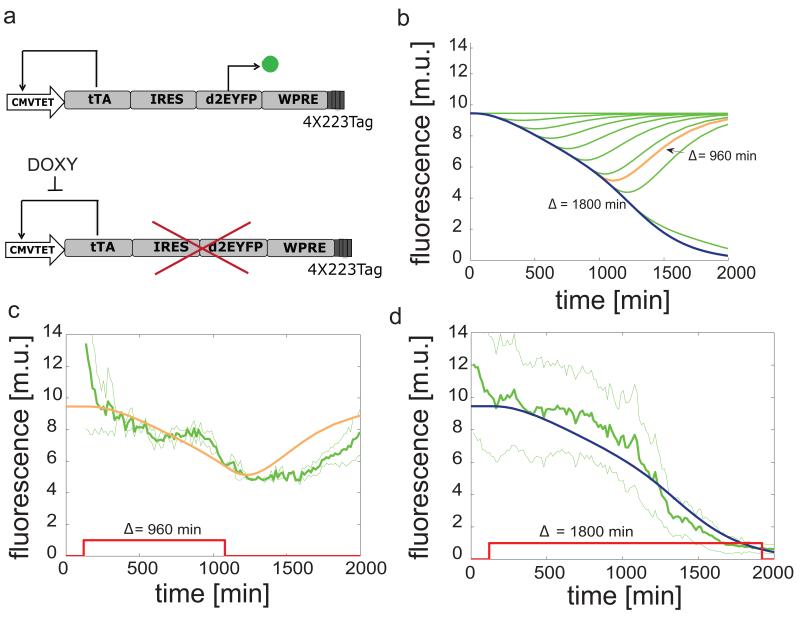
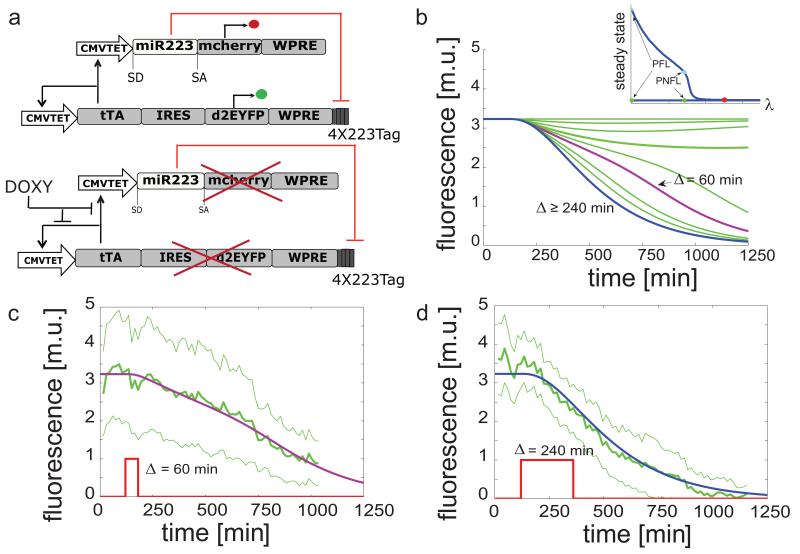
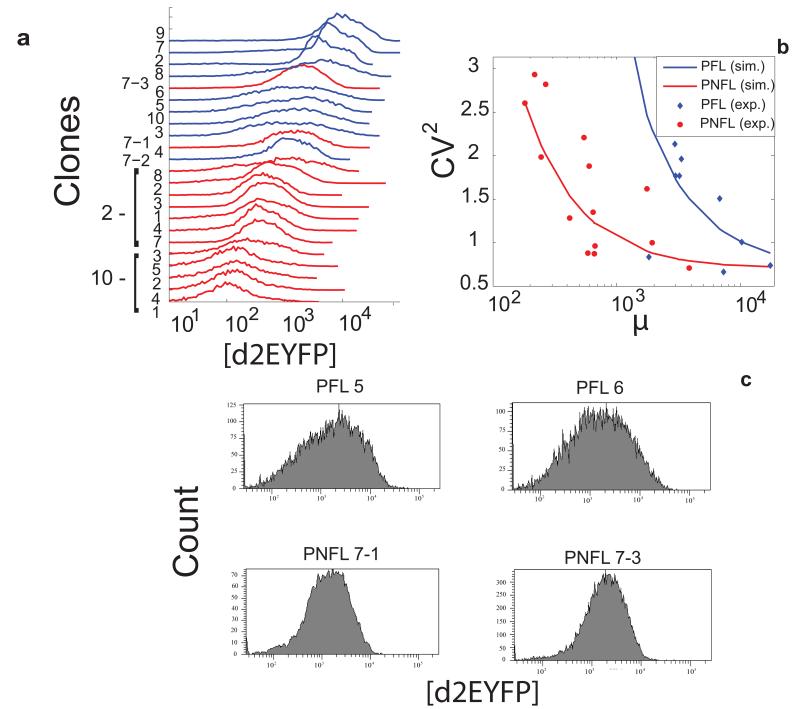
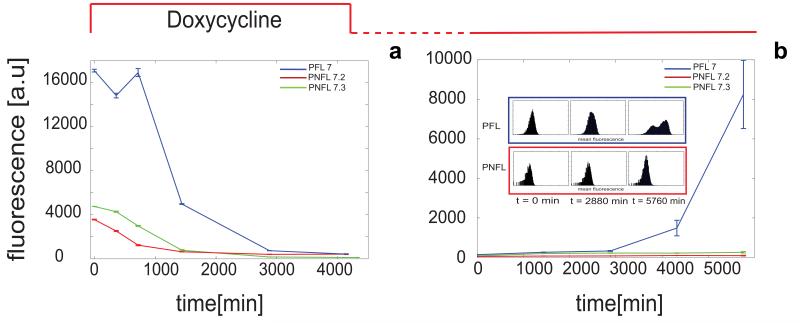
References
-
- Inui M, Martello G, Piccolo S. Microrna control of signal transduction. Nat Rev Mol Cell Biol. 2010;11:264–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
