

A review of phosphate mineral nucleation in biology and geobiology
- PMID: 24077874
- PMCID: PMC3824353
- DOI: 10.1007/s00223-013-9784-9
A review of phosphate mineral nucleation in biology and geobiology
Abstract
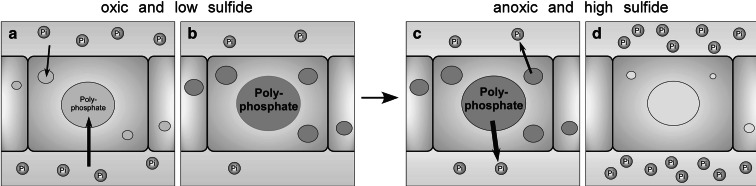
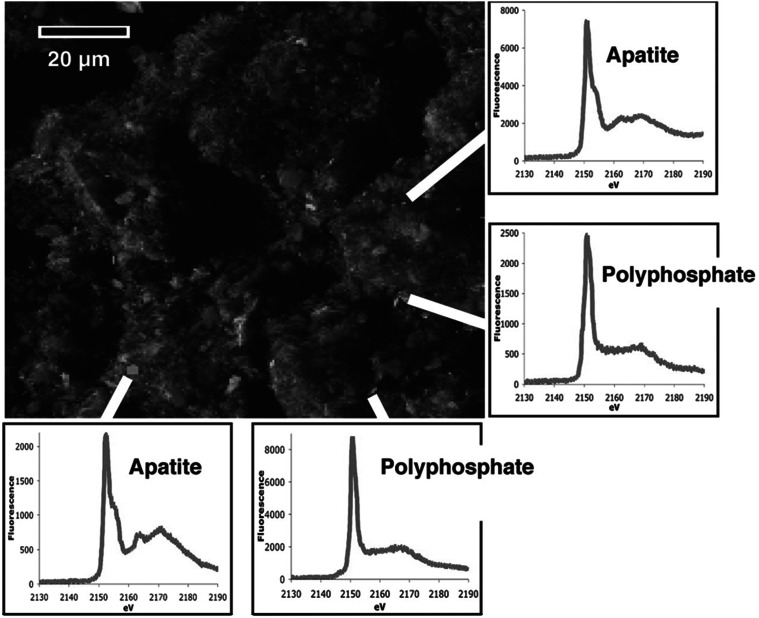
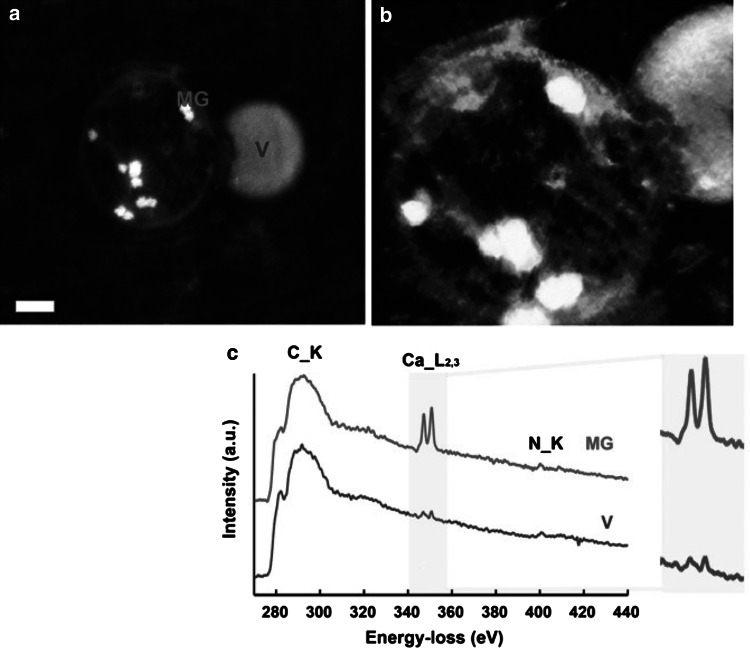
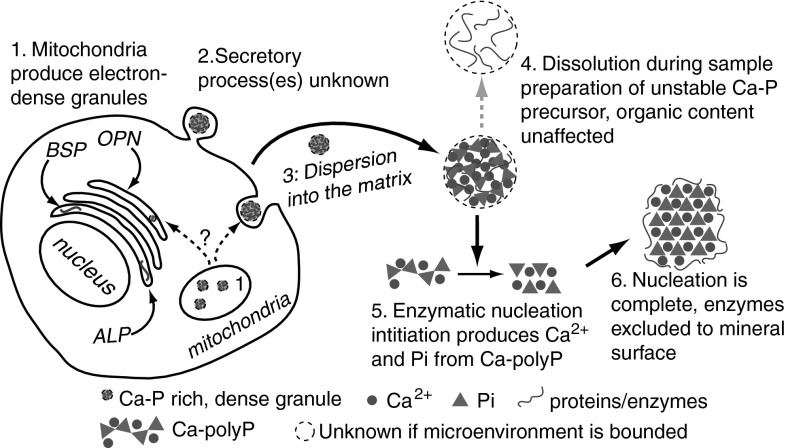
Relationships between geological phosphorite deposition and biological apatite nucleation have often been overlooked. However, similarities in biological apatite and phosphorite mineralogy suggest that their chemical formation mechanisms may be similar. This review serves to draw parallels between two newly described phosphorite mineralization processes, and proposes a similar novel mechanism for biologically controlled apatite mineral nucleation. This mechanism integrates polyphosphate biochemistry with crystal nucleation theory. Recently, the roles of polyphosphates in the nucleation of marine phosphorites were discovered. Marine bacteria and diatoms have been shown to store and concentrate inorganic phosphate (Pi) as amorphous, polyphosphate granules. Subsequent release of these P reserves into the local marine environment as Pi results in biologically induced phosphorite nucleation. Pi storage and release through an intracellular polyphosphate intermediate may also occur in mineralizing oral bacteria. Polyphosphates may be associated with biologically controlled apatite nucleation within vertebrates and invertebrates. Historically, biological apatite nucleation has been attributed to either a biochemical increase in local Pi concentration or matrix-mediated apatite nucleation control. This review proposes a mechanism that integrates both theories. Intracellular and extracellular amorphous granules, rich in both calcium and phosphorus, have been observed in apatite-biomineralizing vertebrates, protists, and atremate brachiopods. These granules may represent stores of calcium-polyphosphate. Not unlike phosphorite nucleation by bacteria and diatoms, polyphosphate depolymerization to Pi would be controlled by phosphatase activity. Enzymatic polyphosphate depolymerization would increase apatite saturation to the level required for mineral nucleation, while matrix proteins would simultaneously control the progression of new biological apatite formation.
Figures
References
-
- Piccoli PM, Candela PA. Apatite in igneous systems. Rev Mineral Geochem. 2002;48:255–292. doi: 10.2138/rmg.2002.48.6. - DOI
-
- Van Cappellen P, Berner RA. A mathematical model for the early diagenesis of phosphorus and fluorine in marine sediments; apatite precipitation. Am J Sci. 1988;288:289–333. doi: 10.2475/ajs.288.4.289. - DOI
-
- Thompson DW. On growth and form. New York: MacMillan; 1945.
-
- Quekett J. On the intimate structure of bone, as composing the skeleton, in the four great classes of animals, viz., mammals, birds, reptiles, and fishes, with some remarks on the great value of the knowledge of such structure in determining the affinities of minute fragments of organic remains. J Microsc. 1849;2:46–58.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
