

Genome-wide profiling reveals stimulus-specific functions of p53 during differentiation and DNA damage of human embryonic stem cells
- PMID: 24078252
- PMCID: PMC3874181
- DOI: 10.1093/nar/gkt866
Genome-wide profiling reveals stimulus-specific functions of p53 during differentiation and DNA damage of human embryonic stem cells
Abstract
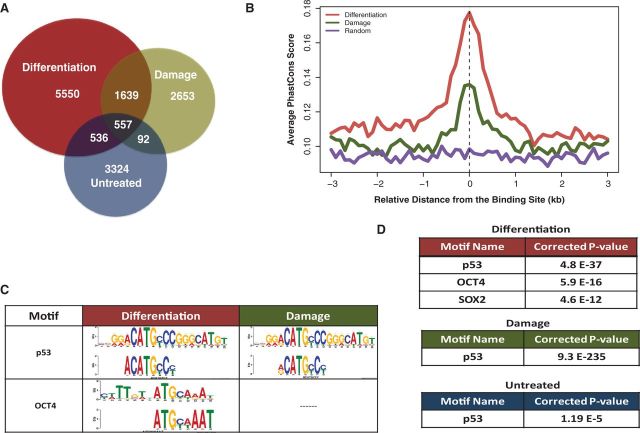
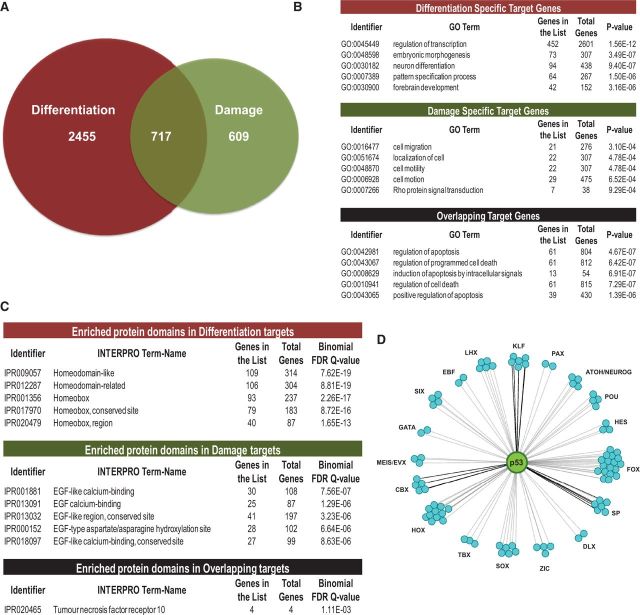
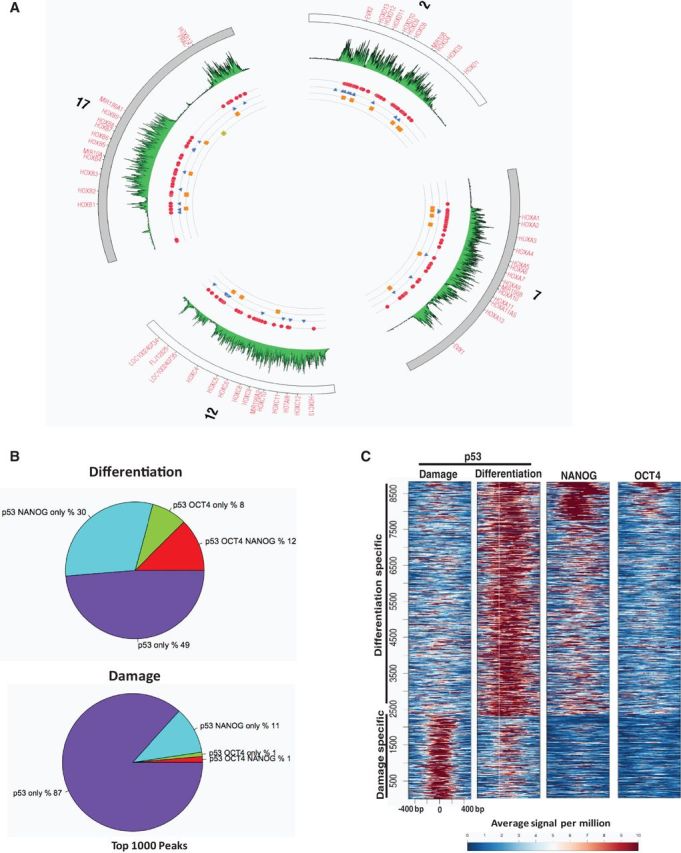
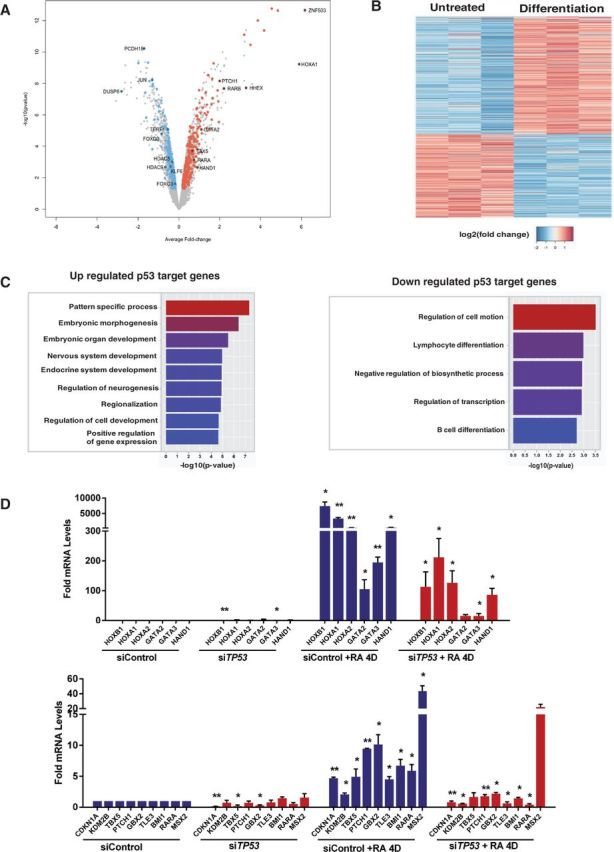
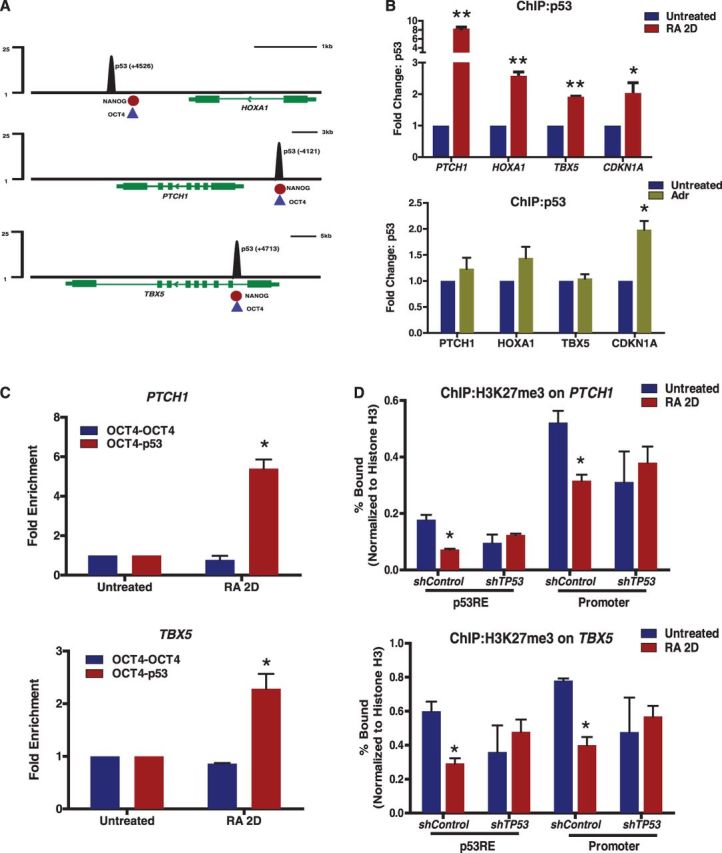
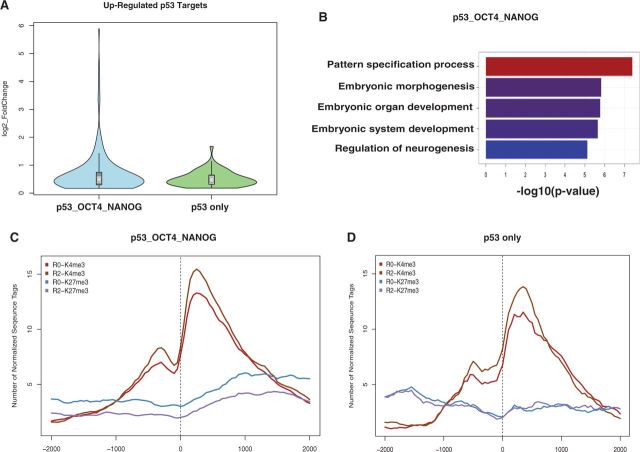
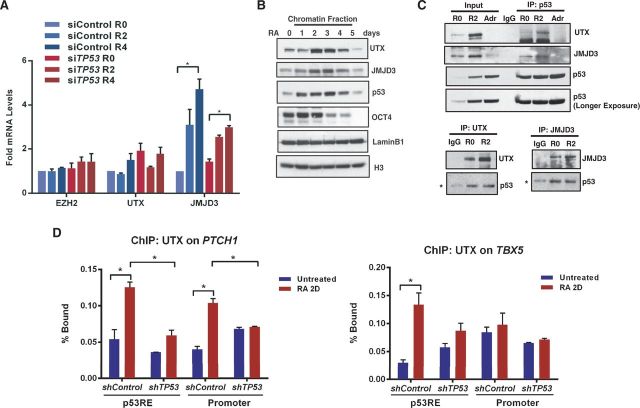
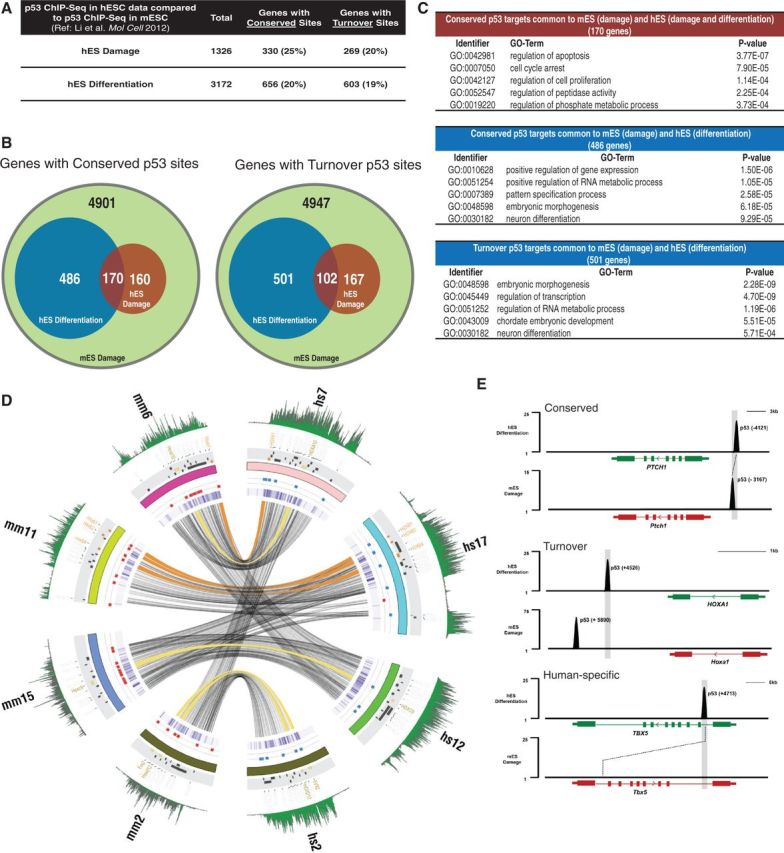
How tumor suppressor p53 selectively responds to specific signals, especially in normal cells, is poorly understood. We performed genome-wide profiling of p53 chromatin interactions and target gene expression in human embryonic stem cells (hESCs) in response to early differentiation, induced by retinoic acid, versus DNA damage, caused by adriamycin. Most p53-binding sites are unique to each state and define stimulus-specific p53 responses in hESCs. Differentiation-activated p53 targets include many developmental transcription factors and, in pluripotent hESCs, are bound by OCT4 and NANOG at chromatin enriched in both H3K27me3 and H3K4me3. Activation of these genes occurs with recruitment of p53 and H3K27me3-specific demethylases, UTX and JMJD3, to chromatin. In contrast, genes associated with cell migration and motility are bound by p53 specifically after DNA damage. Surveillance functions of p53 in cell death and cell cycle regulation are conserved in both DNA damage and differentiation. Comparative genomic analysis of p53-targets in mouse and human ESCs supports an inter-species divergence in p53 regulatory functions during evolution. Our findings expand the registry of p53-regulated genes to define p53-regulated opposition to pluripotency during early differentiation, a process highly distinct from stress-induced p53 response in hESCs.
Figures
References
-
- Vousden KH, Prives C. Blinded by the light: the growing complexity of p53. Cell. 2009;137:413–431. - PubMed
-
- Lin T, Chao C, Saito S, Mazur SJ, Murphy ME, Appella E, Xu Y. p53 induces differentiation of mouse embryonic stem cells by suppressing Nanog expression. Nat. Cell. Biol. 2005;7:165–171. - PubMed
-
- Meletis K, Wirta V, Hede SM, Nister M, Lundeberg J, Frisen J. p53 suppresses the self-renewal of adult neural stem cells. Development. 2006;133:363–369. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
