

Hexokinase-mitochondrial interactions regulate glucose metabolism differentially in adult and neonatal cardiac myocytes
- PMID: 24081983
- PMCID: PMC3787771
- DOI: 10.1085/jgp.201310968
Hexokinase-mitochondrial interactions regulate glucose metabolism differentially in adult and neonatal cardiac myocytes
Abstract
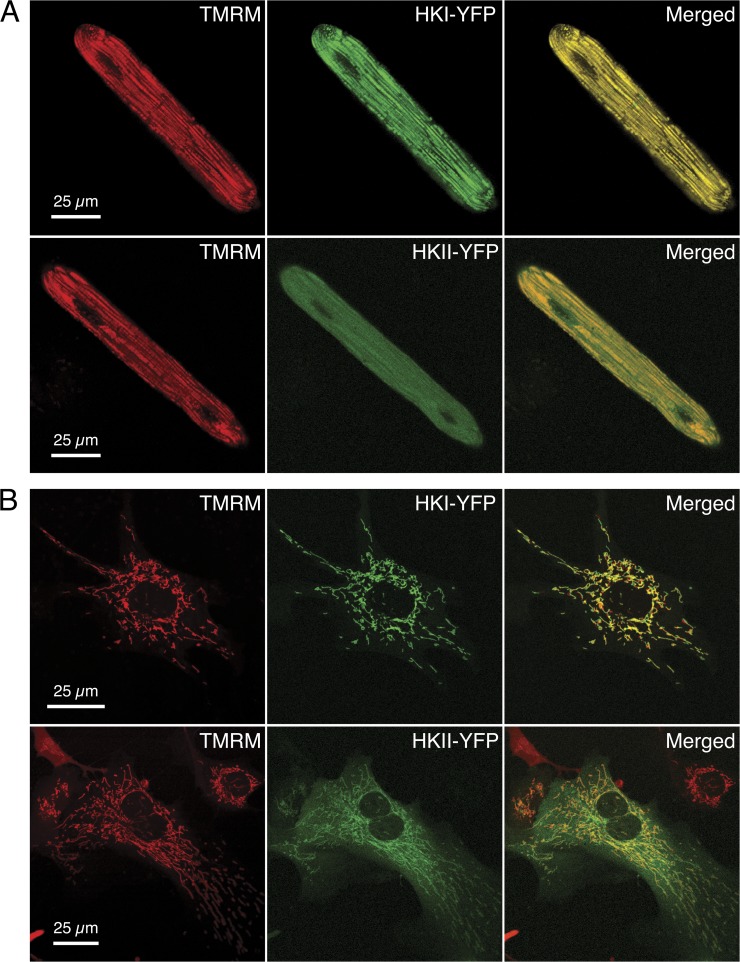
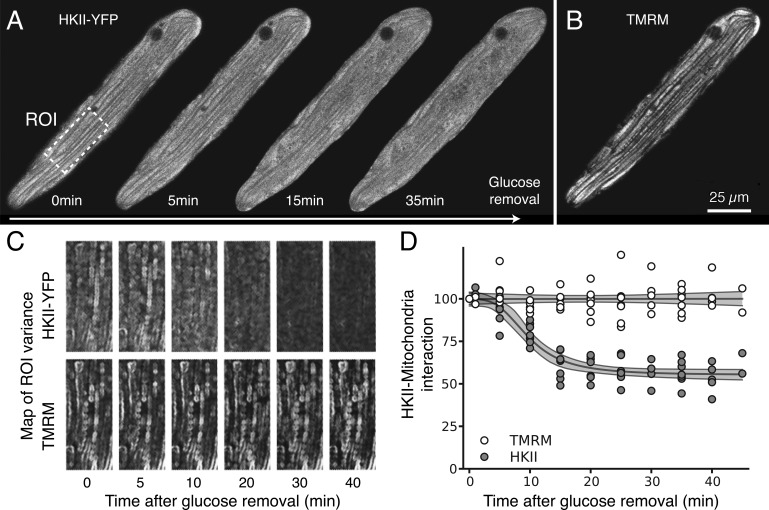
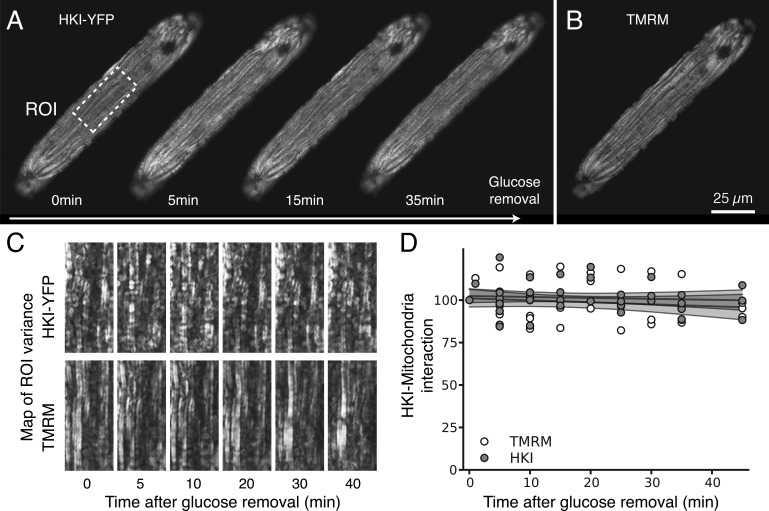
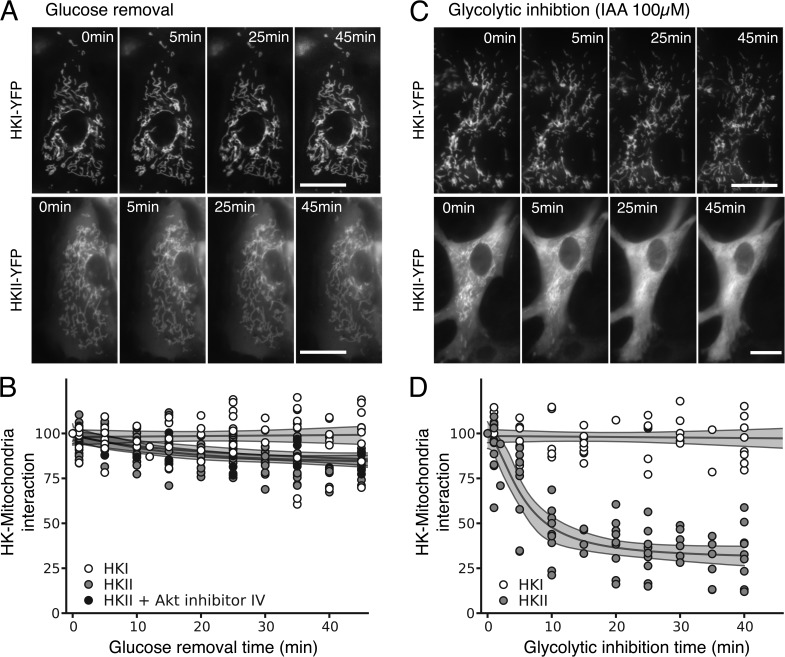
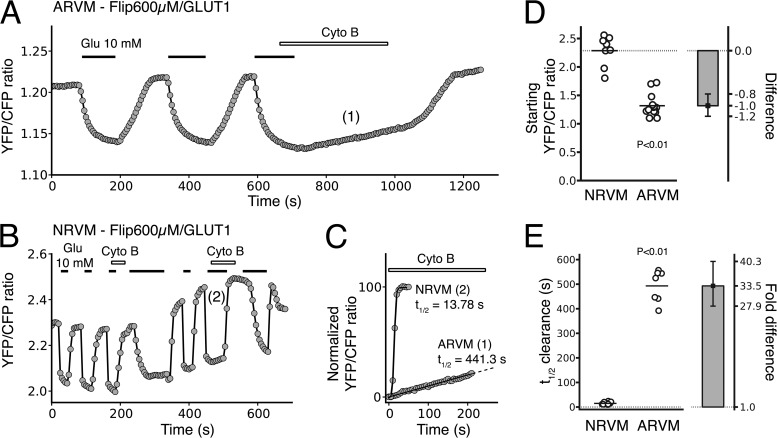
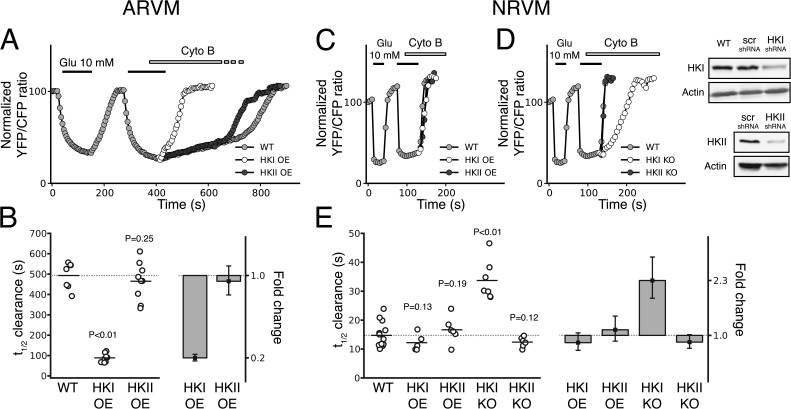
In mammalian tumor cell lines, localization of hexokinase (HK) isoforms to the cytoplasm or mitochondria has been shown to control their anabolic (glycogen synthesis) and catabolic (glycolysis) activities. In this study, we examined whether HK isoform differences could explain the markedly different metabolic profiles between normal adult and neonatal cardiac tissue. We used a set of novel genetically encoded optical imaging tools to track, in real-time in isolated adult (ARVM) and neonatal (NRVM) rat ventricular myocytes, the subcellular distributions of HKI and HKII, and the functional consequences on glucose utilization. We show that HKII, the predominant isoform in ARVM, dynamically translocates from mitochondria and cytoplasm in response to removal of extracellular glucose or addition of iodoacetate (IAA). In contrast, HKI, the predominant isoform in NRVM, is only bound to mitochondria and is not displaced by the above interventions. In ARVM, overexpression of HKI, but not HKII, increased glycolytic activity. In neonatal rat ventricular myocytes (NVRM), knockdown of HKI, but not HKII, decreased glycolytic activity. In conclusion, differential interactions of HKI and HKII with mitochondria underlie the different metabolic profiles of ARVM and NRVM, accounting for the markedly increased glycolytic activity of NRVM.
Figures
References
-
- Aleshin A.E., Zeng C., Bourenkov G.P., Bartunik H.D., Fromm H.J., Honzatko R.B. 1998. The mechanism of regulation of hexokinase: new insights from the crystal structure of recombinant human brain hexokinase complexed with glucose and glucose-6-phosphate. Structure. 6:39–50 10.1016/S0969-2126(98)00006-9 - DOI - PubMed
-
- Burcelin R., Printz R.L., Kande J., Assan R., Granner D.K., Girard J. 1993. Regulation of glucose transporter and hexokinase II expression in tissues of diabetic rats. Am. J. Physiol. 265:E392–E401 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
