

The nodulation factor hydrolase of Medicago truncatula: characterization of an enzyme specifically cleaving rhizobial nodulation signals
- PMID: 24082029
- PMCID: PMC3813642
- DOI: 10.1104/pp.113.223966
The nodulation factor hydrolase of Medicago truncatula: characterization of an enzyme specifically cleaving rhizobial nodulation signals
Abstract
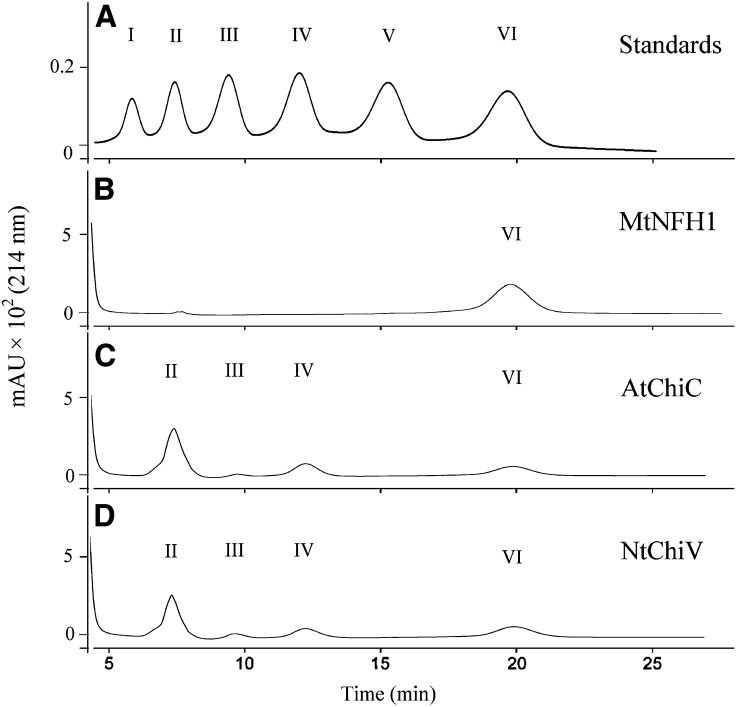
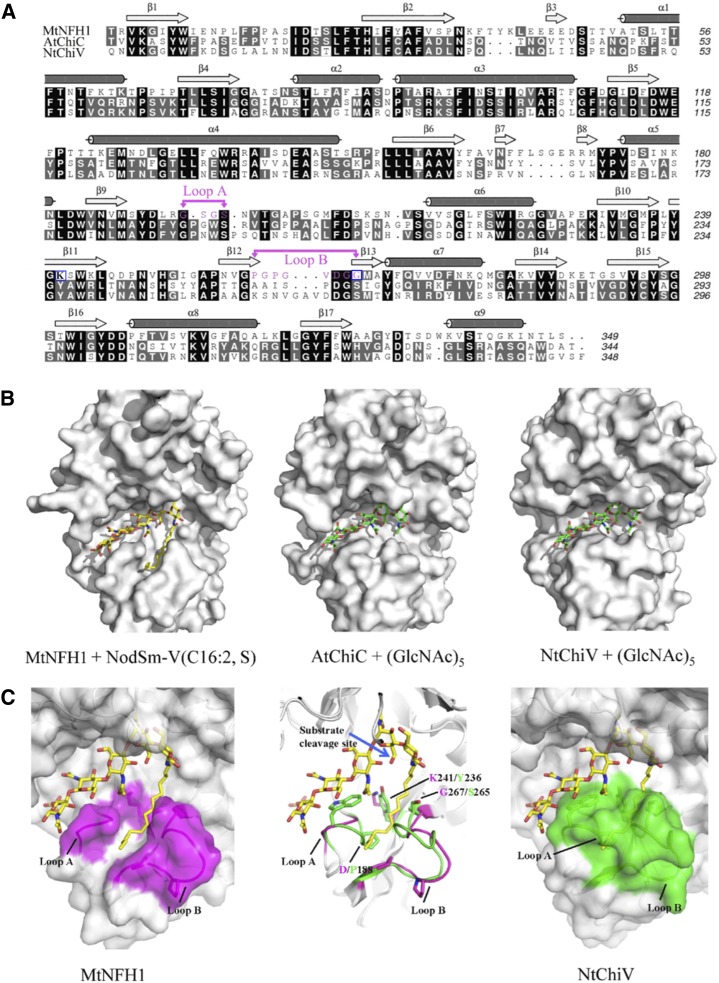
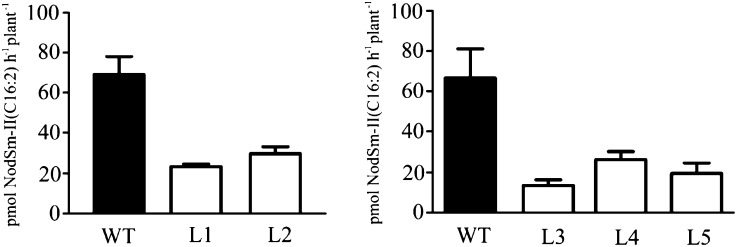
Nodule formation induced by nitrogen-fixing rhizobia depends on bacterial nodulation factors (NFs), modified chitin oligosaccharides with a fatty acid moiety. Certain NFs can be cleaved and inactivated by plant chitinases. However, the most abundant NF of Sinorhizobium meliloti, an O-acetylated and sulfated tetramer, is resistant to hydrolysis by all plant chitinases tested so far. Nevertheless, this NF is rapidly degraded in the host rhizosphere. Here, we identify and characterize MtNFH1 (for Medicago truncatula Nod factor hydrolase 1), a legume enzyme structurally related to defense-related class V chitinases (glycoside hydrolase family 18). MtNFH1 lacks chitinase activity but efficiently hydrolyzes all tested NFs of S. meliloti. The enzyme shows a high cleavage preference, releasing exclusively lipodisaccharides from NFs. Substrate specificity and kinetic properties of MtNFH1 were compared with those of class V chitinases from Arabidopsis (Arabidopsis thaliana) and tobacco (Nicotiana tabacum), which cannot hydrolyze tetrameric NFs of S. meliloti. The Michaelis-Menten constants of MtNFH1 for NFs are in the micromolar concentration range, whereas nonmodified chitin oligosaccharides represent neither substrates nor inhibitors for MtNFH1. The three-dimensional structure of MtNFH1 was modeled on the basis of the known structure of class V chitinases. Docking simulation of NFs to MtNFH1 predicted a distinct binding cleft for the fatty acid moiety, which is absent in the class V chitinases. Point mutation analysis confirmed the modeled NF-MtNFH1 interaction. Silencing of MtNFH1 by RNA interference resulted in reduced NF degradation in the rhizosphere of M. truncatula. In conclusion, we have found a novel legume hydrolase that specifically inactivates NFs.
Figures


References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Antolín-Llovera M, Ried MK, Binder A, Parniske M. (2012) Receptor kinase signaling pathways in plant-microbe interactions. Annu Rev Phytopathol 50: 451–473 - PubMed
-
- Boller T, Gehri A, Mauch F, Vögeli U. (1983) Chitinase in bean leaves: induction by ethylene, purification, properties, and possible function. Planta 157: 22–31 - PubMed
-
- Bradford MM. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
