

Immunology primer for neurosurgeons and neurologists part 2: Innate brain immunity
- PMID: 24083053
- PMCID: PMC3784951
- DOI: 10.4103/2152-7806.118349
Immunology primer for neurosurgeons and neurologists part 2: Innate brain immunity
Abstract
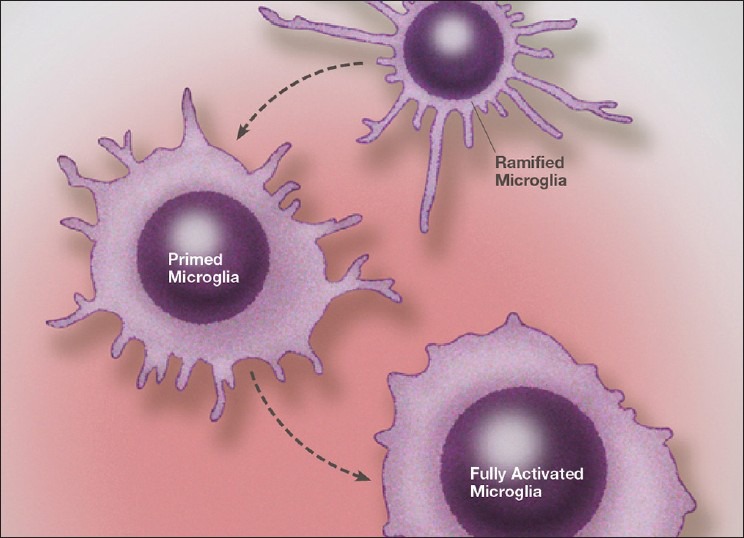
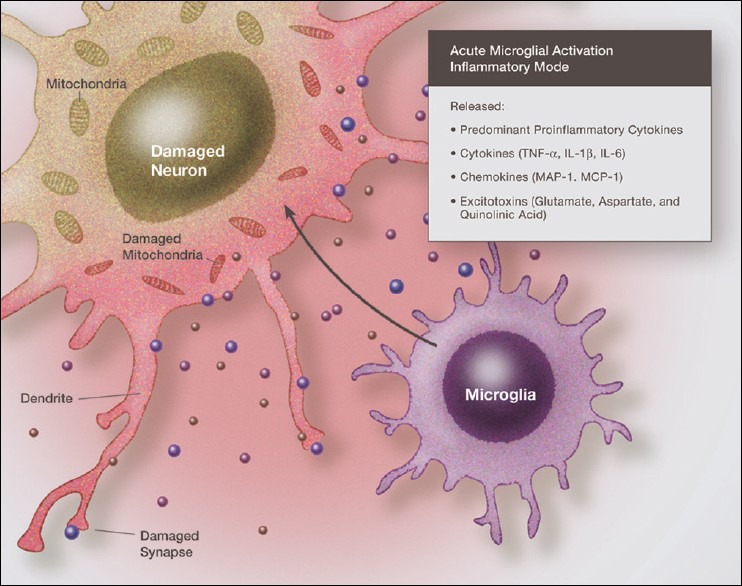
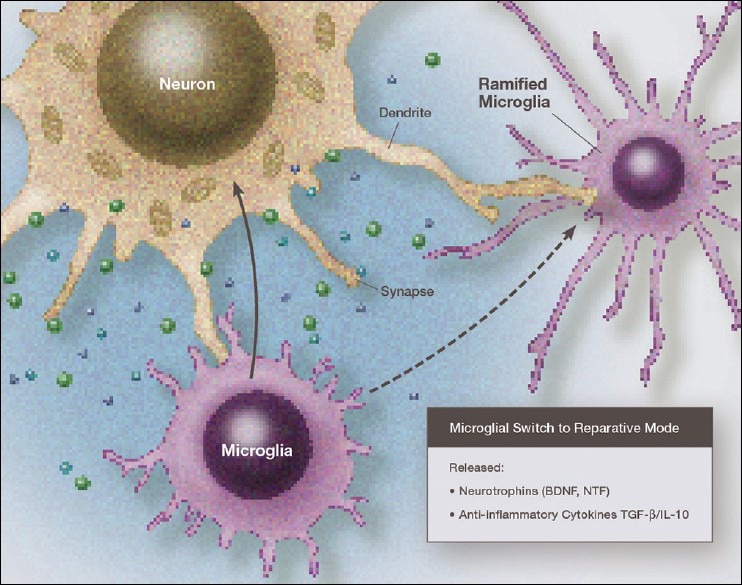
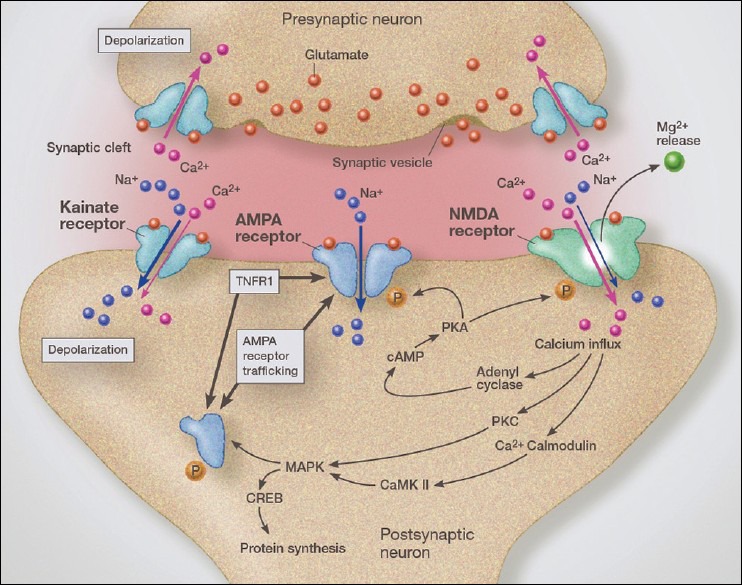
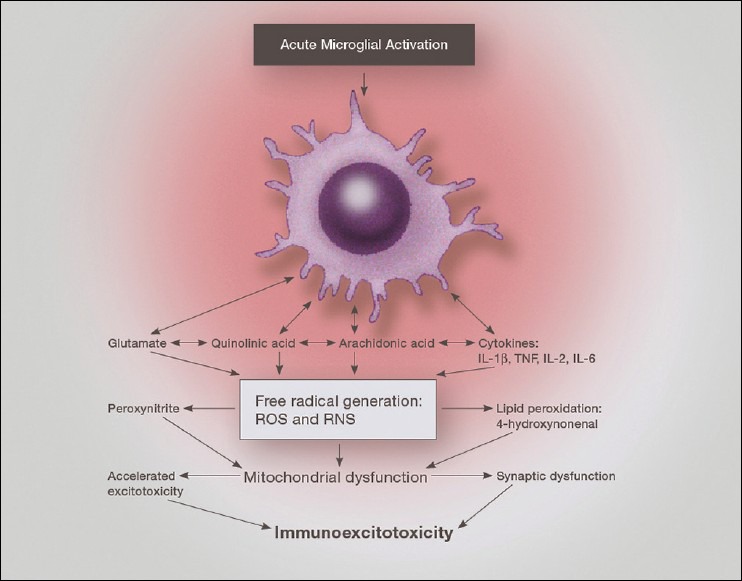
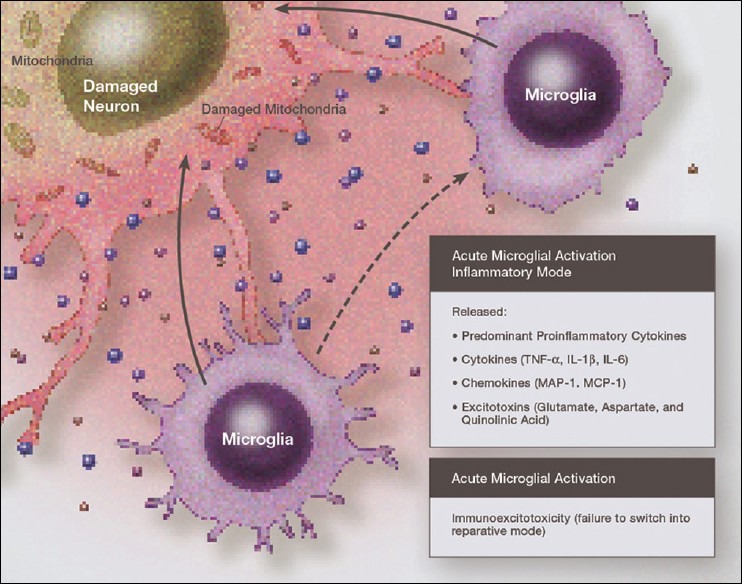
Over the past several decades we have learned a great deal about microglia and innate brain immunity. While microglia are the principle innate immune cells, other cell types also play a role, including invading macrophages, astrocytes, neurons, and endothelial cells. The fastest reacting cell is the microglia and despite its name, resting microglia (also called ramified microglia) are in fact quite active. Motion photomicrographs demonstrate a constant movement of ramified microglial foot processes, which appear to be testing the microenvironment for dangerous alteration in extracellular fluid content. These foot processes, in particular, interact with synapses and play a role in synaptic function. In event of excitatory overactivity, these foot processes can strip selected synapses, thus reducing activation states as a neuroprotective mechanism. They can also clear extracellular glutamate so as to reduce the risk of excitotoxicity. Microglia also appear to have a number of activation phenotypes, such as: (1) phagocytic, (2) neuroprotective and growth promoting, or (3) primarily neurodestructive. These innate immune cells can migrate a great distance under pathological conditions and appear to have anatomic specificity, meaning they can accumulate in specifically selected areas of the brain. There is some evidence that there are several types of microglia. Macrophage infiltration into the embryonic brain is the source of resident microglia and in adulthood macrophages can infiltrate the brain and are for the most part pathologically indistinguishable from resident microglia, but may react differently. Activation itself does not imply a destructive phenotype and can be mostly neuroprotective via phagocytosis of debris, neuron parts and dying cells and by the release of neurotrophins such as nerve growth factor (NGF) and brain derived neurotrophic factor (BDNF). Evidence is accumulating that microglia undergo dynamic fluctuations in phenotype as the neuropathology evolves. For example, in the early stages of neurotrauma and stroke, microglia play a mostly neuroprotective role and only later switch to a neurodestructive mode. A great number of biological systems alter microglia function, including neurohormones, cannabinoids, other neurotransmitters, adenosine triphosphate (ATP), adenosine, and corticosteroids. One can appreciate that with aging many of these systems are altered by the aging process itself or by disease thus changing the sensitivity of the innate immune system.
Keywords: Immune surface receptors; immunoexcitotoxicity; innate immunity; microglia; microglial priming.
Figures
References
-
- Akira S, Takeda K. Toll-like receptor signaling. Nat Rev Immunol. 2004;4:499–511. - PubMed
-
- Alarcon R, Fuenzalida C, Santibanez M, von Bernhardt R. Expression of scavenger receptors in glial cells. J Biol Chem. 2005;280:30406–15. - PubMed
-
- Aloisi F, Ria F, Adorini L. Regulation of T-cell responses by CNS antigen-presenting cell: Different roles for microglia and astrocytes. Immunol Today. 2000;21:141–7. - PubMed
-
- Aloisi F, Ria F, Penna G, Adorini L. Microglia are more efficient than astrocytes in antigen processing and in the Th1 but not Th2 activation. J Immunol. 1998;160:4671–80. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
