

Control of mRNA stability contributes to low levels of nuclear poly(A) binding protein 1 (PABPN1) in skeletal muscle
- PMID: 24083404
- PMCID: PMC3879409
- DOI: 10.1186/2044-5040-3-23
Control of mRNA stability contributes to low levels of nuclear poly(A) binding protein 1 (PABPN1) in skeletal muscle
Abstract
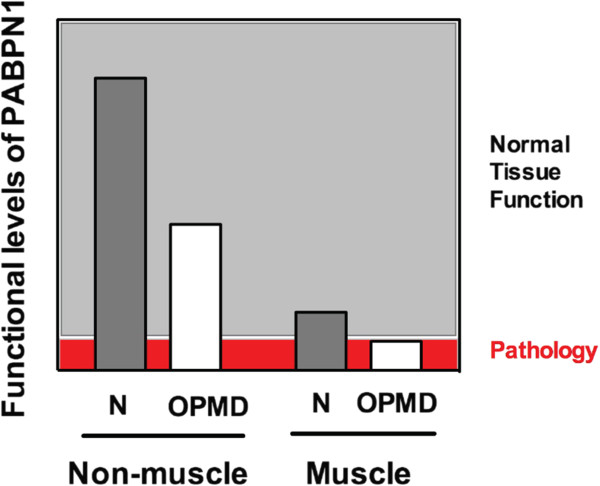
Background: The nuclear poly(A) binding protein 1 (PABPN1) is a ubiquitously expressed protein that plays critical roles at multiple steps in post-transcriptional regulation of gene expression. Short expansions of the polyalanine tract in the N-terminus of PABPN1 lead to oculopharyngeal muscular dystrophy (OPMD), which is an adult onset disease characterized by eyelid drooping, difficulty in swallowing, and weakness in the proximal limb muscles. Why alanine-expanded PABPN1 leads to muscle-specific pathology is unknown. Given the general function of PABPN1 in RNA metabolism, intrinsic characteristics of skeletal muscle may make this tissue susceptible to the effects of mutant PABPN1.
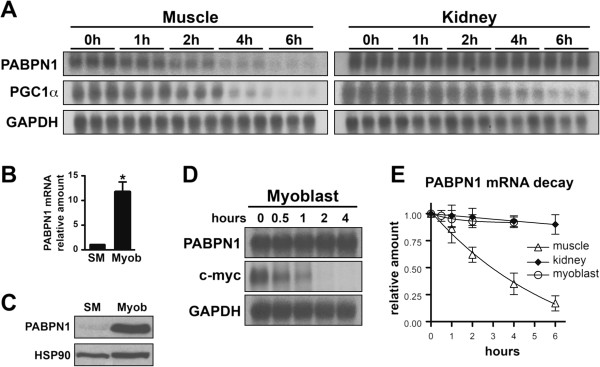
Methods: To begin to understand the muscle specificity of OPMD, we investigated the steady-state levels of PABPN1 in different tissues of humans and mice. Additionally, we analyzed the levels of PABPN1 during muscle regeneration after injury in mice. Furthermore, we assessed the dynamics of PABPN1 mRNA decay in skeletal muscle compared to kidney.
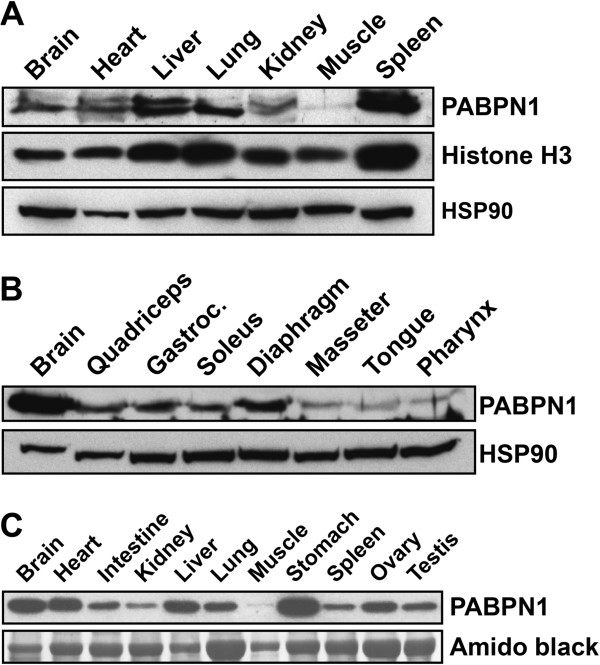
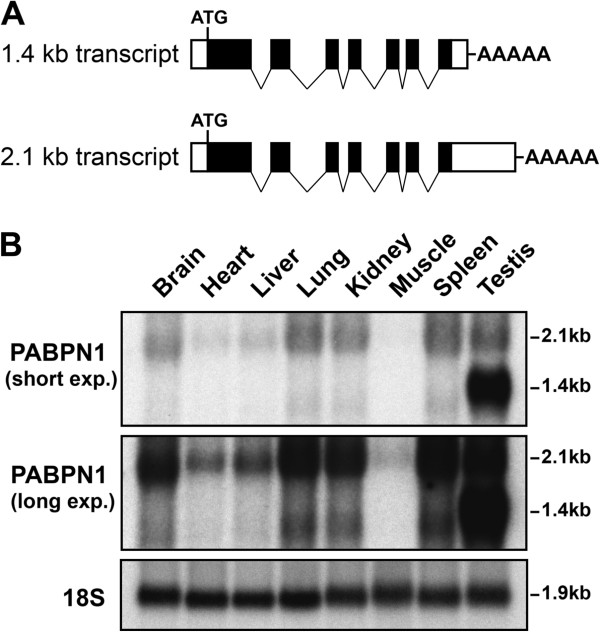
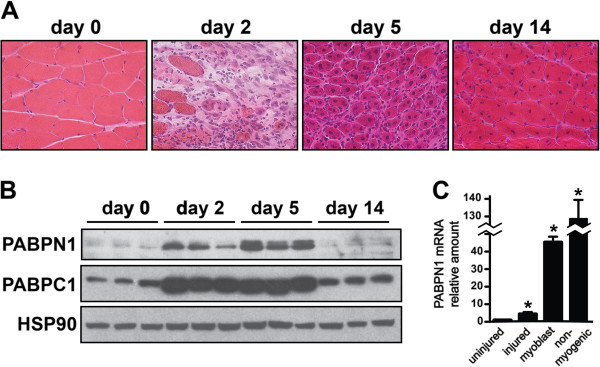
Results: Here, we show that the steady-state levels of both PABPN1 mRNA and protein are drastically lower in mouse and human skeletal muscle, particularly those impacted in OPMD, compared to other tissues. In contrast, PABPN1 levels are increased during muscle regeneration, suggesting a greater requirement for PABPN1 function during tissue repair. Further analysis indicates that modulation of PABPN1 expression is likely due to post-transcriptional mechanisms acting at the level of mRNA stability.
Conclusions: Our results demonstrate that PABPN1 steady-state levels and likely control of expression differ significantly in skeletal muscle as compared to other tissues, which could have important implications for understanding the muscle-specific nature of OPMD.
Figures
References
-
- Kuhn U, Gundel M, Knoth A, Kerwitz Y, Rudel S, Wahle E. Poly(A) tail length is controlled by the nuclear poly(A)-binding protein regulating the interaction between poly(A) polymerase and the cleavage and polyadenylation specificity factor. J Biol Chem. 2009;3:22803–22814. doi: 10.1074/jbc.M109.018226. - DOI - PMC - PubMed
-
- Jenal M, Elkon R, Loayza-Puch F, van Haaften G, Kühn U, Menzies FM, Oude Vrielink JA, Bos AJ, Drost J, Rooijers K, Rubinsztein DC, Agami R. The poly(A)-binding protein nuclear 1 suppresses alternative cleavage and polyadenylation sites. Cell. 2012;3:538–553. doi: 10.1016/j.cell.2012.03.022. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
