

Embryonic heart progenitors and cardiogenesis
- PMID: 24086063
- PMCID: PMC3784811
- DOI: 10.1101/cshperspect.a013847
Embryonic heart progenitors and cardiogenesis
Abstract
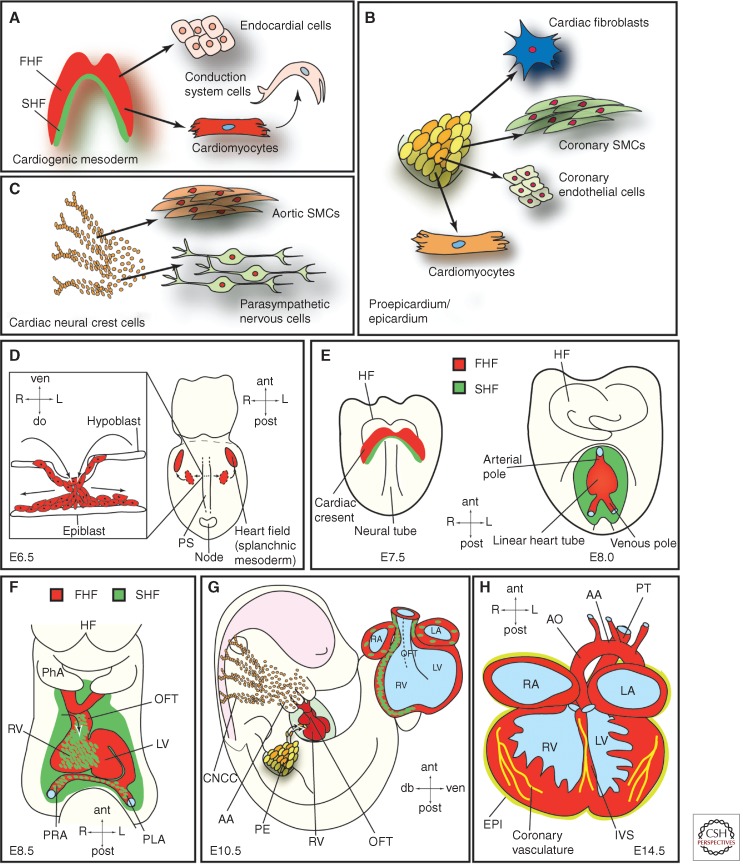
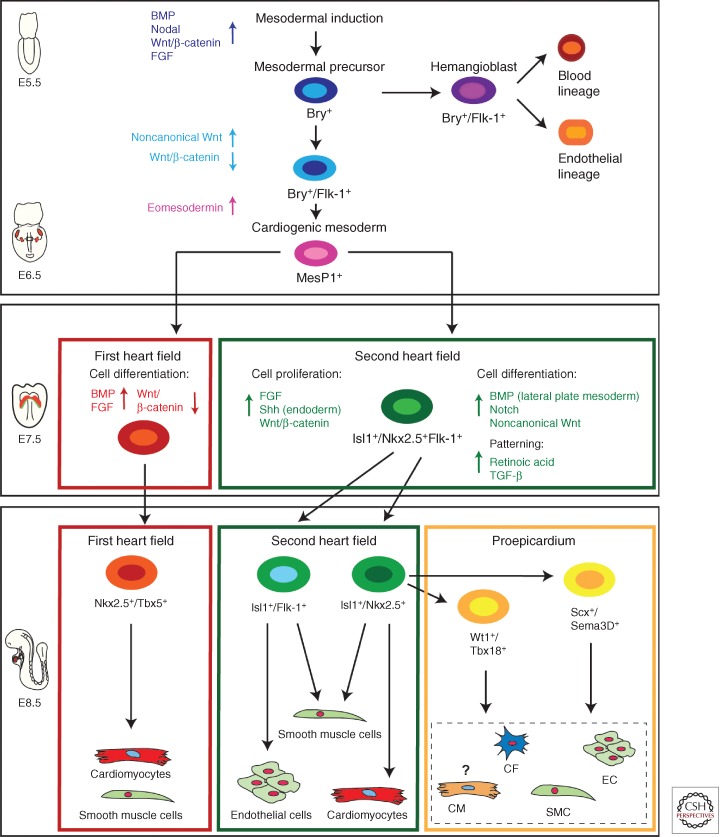
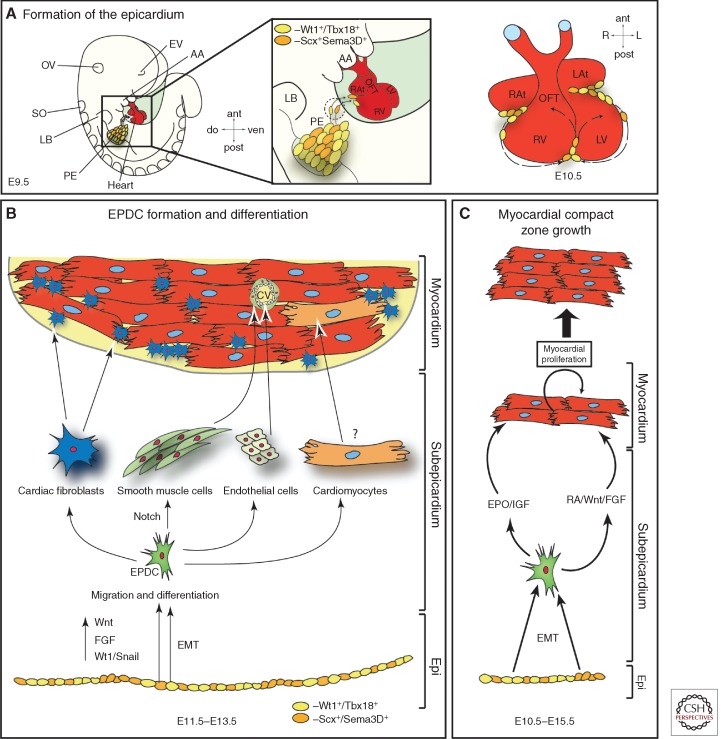
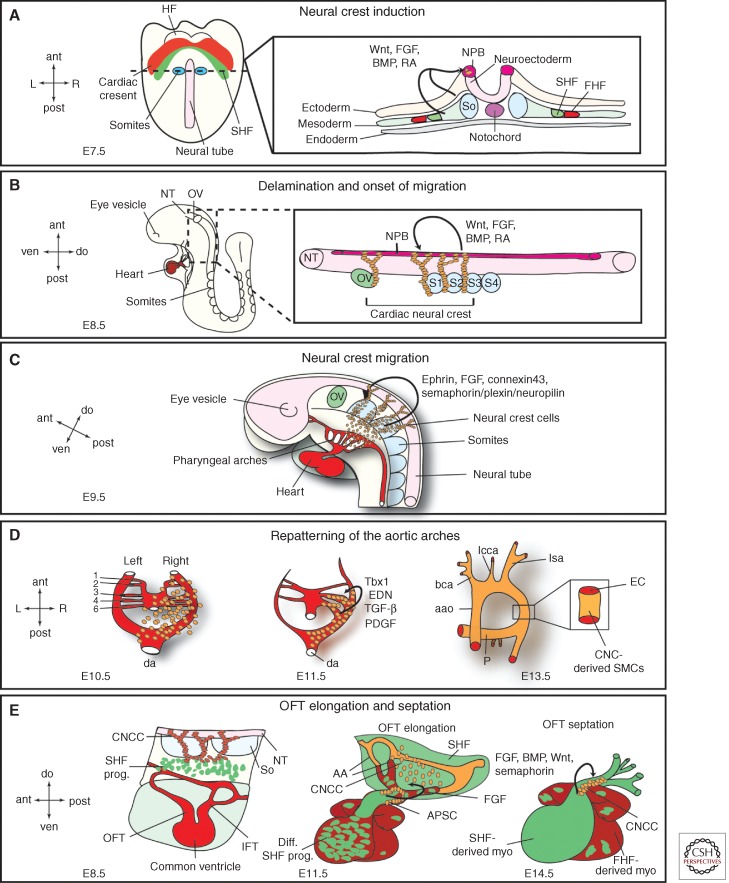
The mammalian heart is a highly specialized organ, comprised of many different cell types arising from distinct embryonic progenitor populations during cardiogenesis. Three precursor populations have been identified to contribute to different myocytic and nonmyocytic cell lineages of the heart: cardiogenic mesoderm cells (CMC), the proepicardium (PE), and cardiac neural crest cells (CNCCs). This review will focus on molecular cues necessary for proper induction, expansion, and lineage-specific differentiation of these progenitor populations during cardiac development in vivo. Moreover, we will briefly discuss how the knowledge gained on embryonic heart progenitor biology can be used to develop novel therapeutic strategies for the management of congenital heart disease as well as for improvement of cardiac function in ischemic heart disease.
Figures
References
-
- Abu-Issa R, Smyth G, Smoak I, Yamamura K, Meyers EN 2002. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development 129: 4613–4625 - PubMed
-
- Abu-Issa R, Waldo K, Kirby ML 2004. Heart fields: One, two or more? Dev Biol 272: 281–285 - PubMed
-
- Aguirre A, Sancho-Martinez I, Izpisua Belmonte JC 2013. Reprogramming toward heart regeneration: Stem cells and beyond. Cell Stem Cell 12: 275–284 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources