

Hypoxic regulation of hand1 controls the fetal-neonatal switch in cardiac metabolism
- PMID: 24086110
- PMCID: PMC3782421
- DOI: 10.1371/journal.pbio.1001666
Hypoxic regulation of hand1 controls the fetal-neonatal switch in cardiac metabolism
Erratum in
- PLoS Biol. 2013 Dec;11(12). doi:10.1371/annotation/a9a7f37a-3fa7-4f7f-8310-1339bf5a666e
Abstract
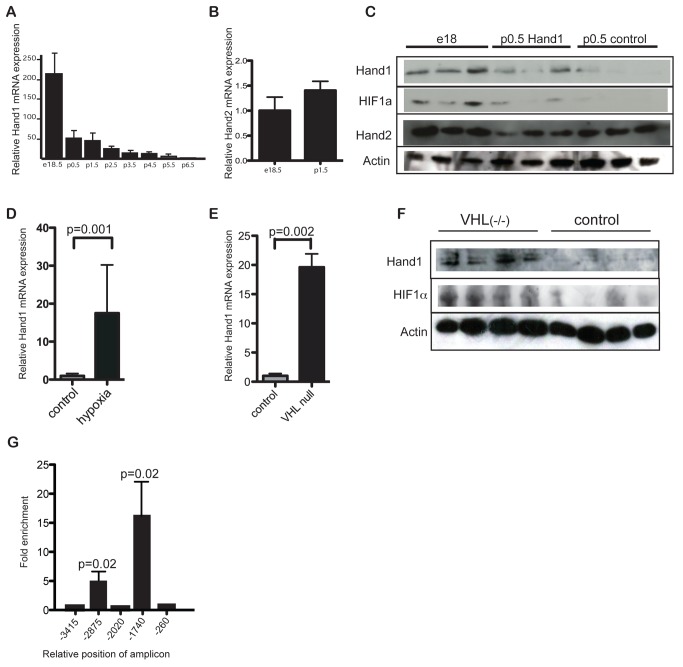
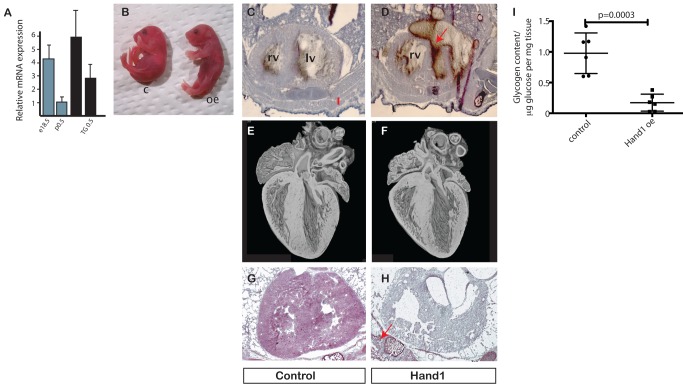
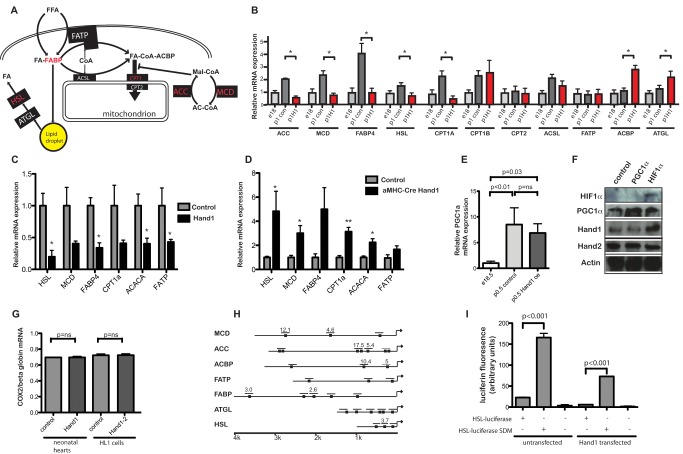
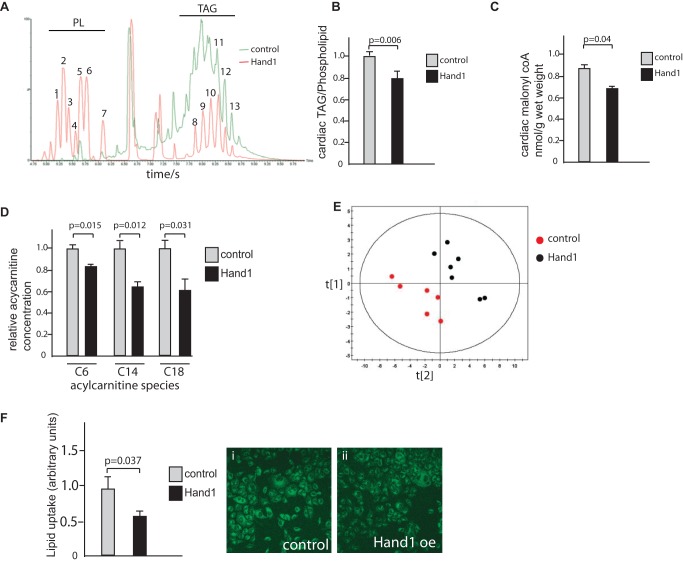
Cardiomyocytes are vulnerable to hypoxia in the adult, but adapted to hypoxia in utero. Current understanding of endogenous cardiac oxygen sensing pathways is limited. Myocardial oxygen consumption is determined by regulation of energy metabolism, which shifts from glycolysis to lipid oxidation soon after birth, and is reversed in failing adult hearts, accompanying re-expression of several "fetal" genes whose role in disease phenotypes remains unknown. Here we show that hypoxia-controlled expression of the transcription factor Hand1 determines oxygen consumption by inhibition of lipid metabolism in the fetal and adult cardiomyocyte, leading to downregulation of mitochondrial energy generation. Hand1 is under direct transcriptional control by HIF1α. Transgenic mice prolonging cardiac Hand1 expression die immediately following birth, failing to activate the neonatal lipid metabolising gene expression programme. Deletion of Hand1 in embryonic cardiomyocytes results in premature expression of these genes. Using metabolic flux analysis, we show that Hand1 expression controls cardiomyocyte oxygen consumption by direct transcriptional repression of lipid metabolising genes. This leads, in turn, to increased production of lactate from glucose, decreased lipid oxidation, reduced inner mitochondrial membrane potential, and mitochondrial ATP generation. We found that this pathway is active in adult cardiomyocytes. Up-regulation of Hand1 is protective in a mouse model of myocardial ischaemia. We propose that Hand1 is part of a novel regulatory pathway linking cardiac oxygen levels with oxygen consumption. Understanding hypoxia adaptation in the fetal heart may allow development of strategies to protect cardiomyocytes vulnerable to ischaemia, for example during cardiac ischaemia or surgery.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Fine-tuning metabolic switches.PLoS Biol. 2013 Sep;11(9):e1001664. doi: 10.1371/journal.pbio.1001664. Epub 2013 Sep 24. PLoS Biol. 2013. PMID: 24086108 Free PMC article. No abstract available.
References
-
- Lopaschuk GD, Spafford MA, Marsh DR (1991) Glycolysis is predominant source of myocardial ATP production immediately after birth. Am J Physiol 261: H1698–1705. - PubMed
-
- Lopaschuk GD, Jaswal JS (2010) Energy metabolic phenotype of the cardiomyocyte during development, differentiation, and postnatal maturation. J Cardiovasc Pharmacol 56: 130–140. - PubMed
-
- Sheldon CA, Friedman WF, Sybers HD (1976) Scanning electron microscopy of fetal and neonatal lamb cardiac cells. J Mol Cell Cardiol 8: 853–862. - PubMed
-
- Fisher DJ, Heymann MA, Rudolph AM (1980) Myocardial oxygen and carbohydrate consumption in fetal lambs in utero and in adult sheep. Am J Physiol 238: H399–405. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
