

Febuxostat, an inhibitor of xanthine oxidase, suppresses lipopolysaccharide-induced MCP-1 production via MAPK phosphatase-1-mediated inactivation of JNK
- PMID: 24086554
- PMCID: PMC3783396
- DOI: 10.1371/journal.pone.0075527
Febuxostat, an inhibitor of xanthine oxidase, suppresses lipopolysaccharide-induced MCP-1 production via MAPK phosphatase-1-mediated inactivation of JNK
Abstract
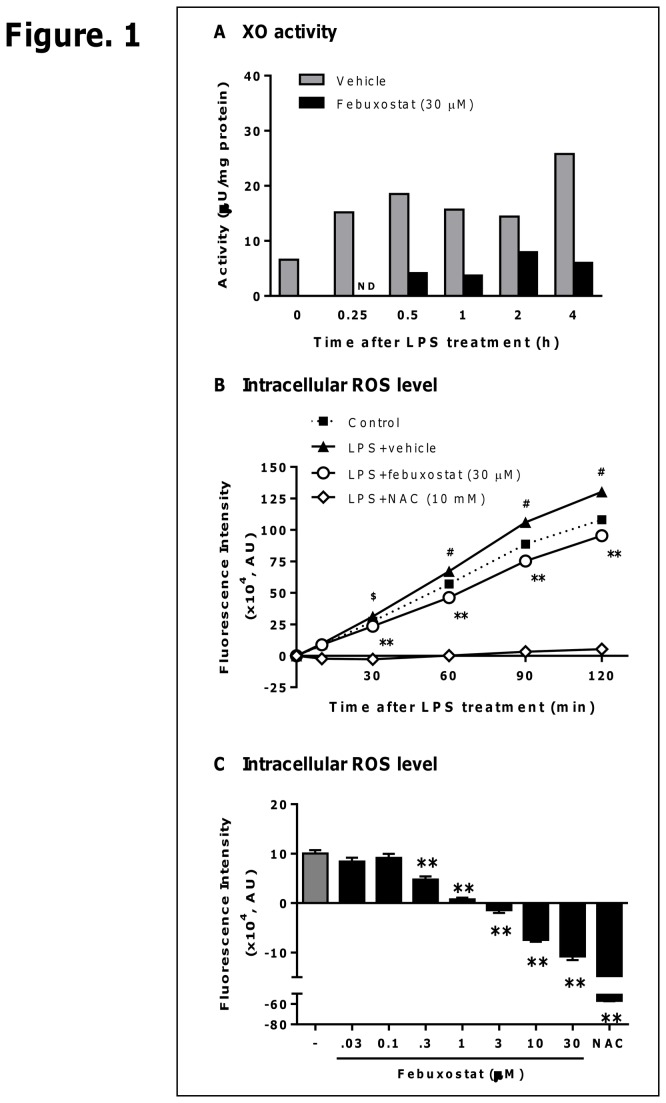
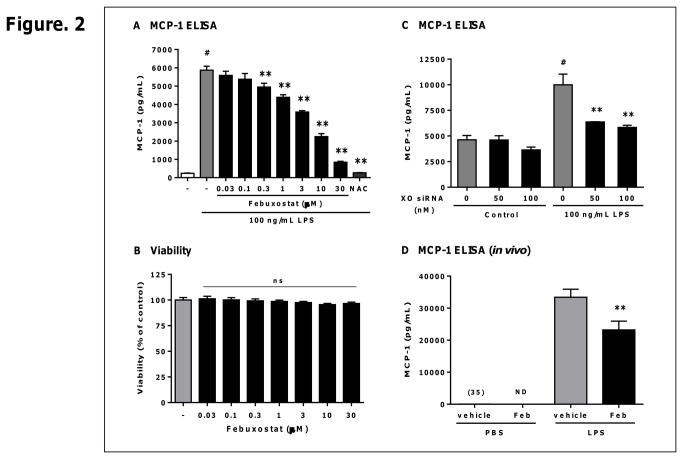
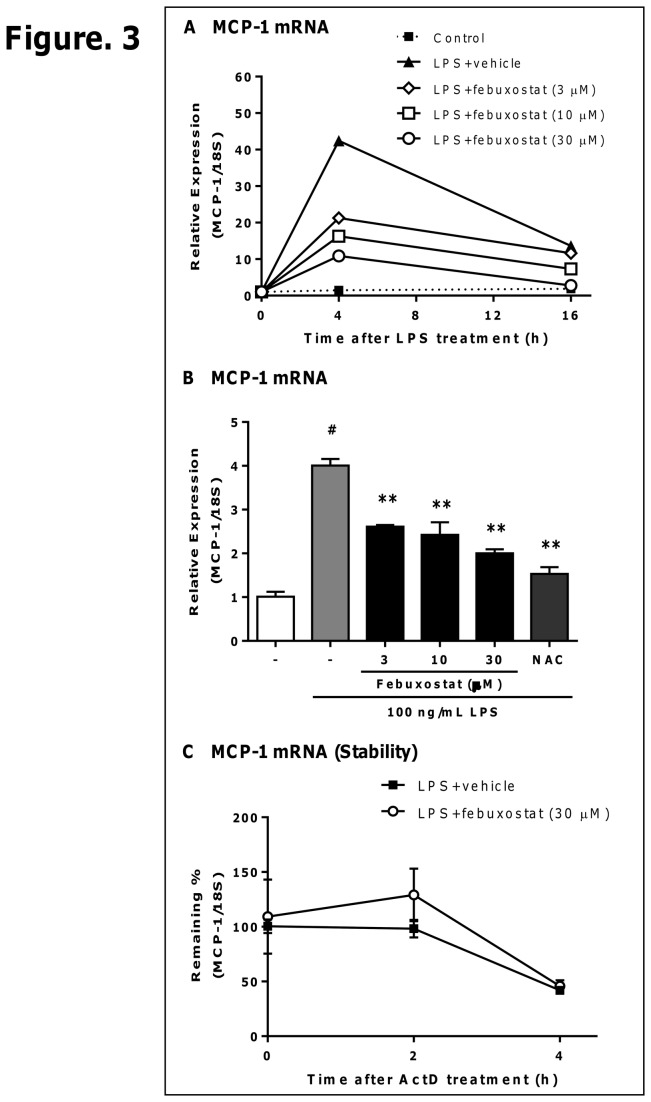
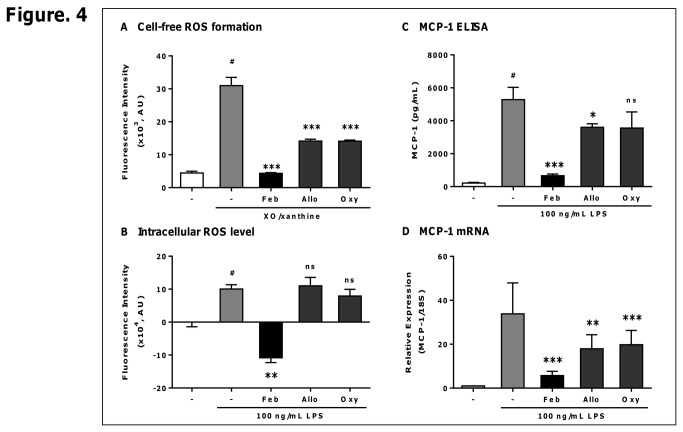
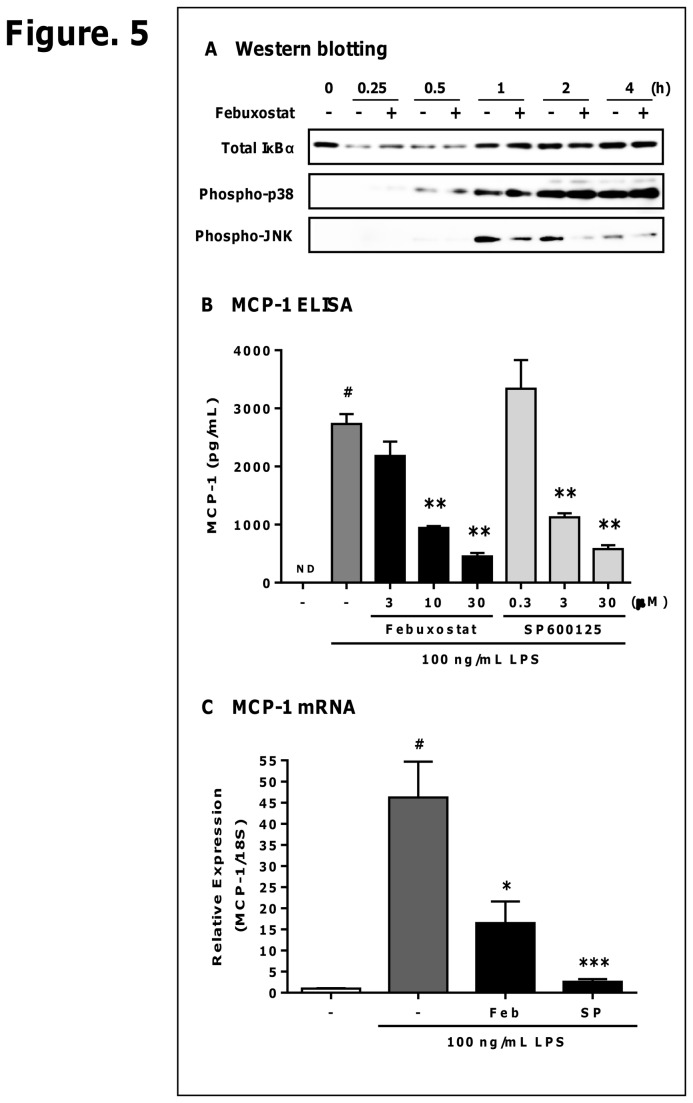
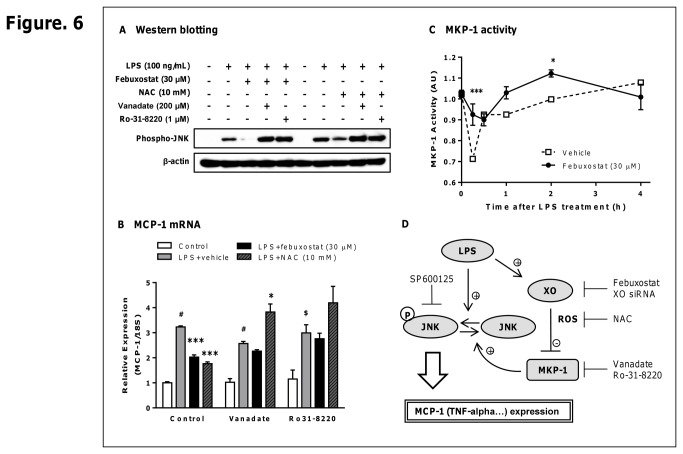
Excess reactive oxygen species (ROS) formation can trigger various pathological conditions such as inflammation, in which xanthine oxidase (XO) is one major enzymatic source of ROS. Although XO has been reported to play essential roles in inflammatory conditions, the molecular mechanisms underlying the involvement of XO in inflammatory pathways remain unclear. Febuxostat, a selective and potent inhibitor of XO, effectively inhibits not only the generation of uric acid but also the formation of ROS. In this study, therefore, we examined the effects of febuxostat on lipopolysaccharide (LPS)-mediated inflammatory responses. Here we show that febuxostat suppresses LPS-induced MCP-1 production and mRNA expression via activating MAPK phosphatase-1 (MKP-1) which, in turn, leads to dephosphorylation and inactivation of JNK in macrophages. Moreover, these effects of febuxostat are mediated by inhibiting XO-mediated intracellular ROS production. Taken together, our data suggest that XO mediates LPS-induced phosphorylation of JNK through ROS production and MKP-1 inactivation, leading to MCP-1 production in macrophages. These studies may bring new insights into the novel role of XO in regulating inflammatory process through MAPK phosphatase, and demonstrate the potential use of XO inhibitor in modulating the inflammatory processes.
Conflict of interest statement
Figures
References
-
- Park JG, Oh GT (2011) The role of peroxidases in the pathogenesis of atherosclerosis. BMB Rep 44: 497-505. doi:10.5483/BMBRep.2011.44.8.497. PubMed: 21871172. - DOI - PubMed
-
- Filippin LI, Vercelino R, Marroni NP, Xavier RM (2008) Redox signalling and the inflammatory response in rheumatoid arthritis. Clin Exp Immunol 152: 415-422. doi:10.1111/j.1365-2249.2008.03634.x. PubMed: 18422737. - DOI - PMC - PubMed
-
- Cape JL, Hurst JK (2009) The role of nitrite ion in phagocyte function--perspectives and puzzles. Arch Biochem Biophys 484: 190-196. doi:10.1016/j.abb.2009.01.010. PubMed: 19402211. - DOI - PMC - PubMed
-
- Berry CE, Hare JM (2004) Xanthine oxidoreductase and cardiovascular disease: molecular mechanisms and pathophysiological implications. J Physiol 555: 589-606. doi:10.1113/jphysiol.2003.055913. PubMed: 14694147. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
