

Anti-HIV activity of human defensin 5 in primary CD4+ T cells under serum-deprived conditions is a consequence of defensin-mediated cytotoxicity
- PMID: 24086683
- PMCID: PMC3783372
- DOI: 10.1371/journal.pone.0076038
Anti-HIV activity of human defensin 5 in primary CD4+ T cells under serum-deprived conditions is a consequence of defensin-mediated cytotoxicity
Abstract
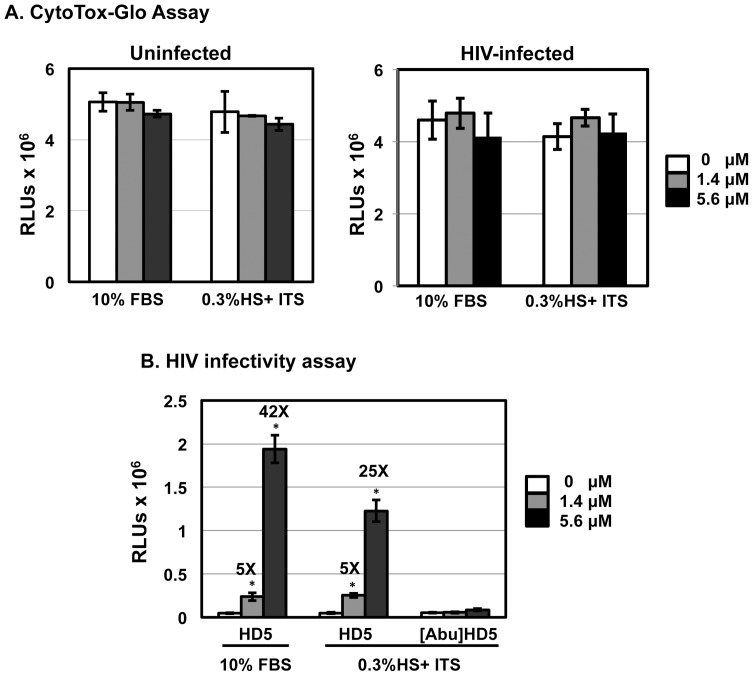
Background: We have previously shown that human defensin 5 (HD5) promotes HIV infectivity in both primary CD4+ T cells and HeLa cells expressing CD4 and CCR5. HD5 is induced in response to sexually transmitted infections (STIs) such as Chlamydia trachomatis and Neisseria gonorrhoeae, suggesting it plays a role in STI-mediated enhancement of HIV transmission. In contrast to our findings, a recent study reports that HD5 has an anti-HIV effect in primary CD4+ T cells under serum-deprived conditions. To resolve these apparently contradictory observations, we investigated experimental parameters that might contribute to contrasting effects of HD5.
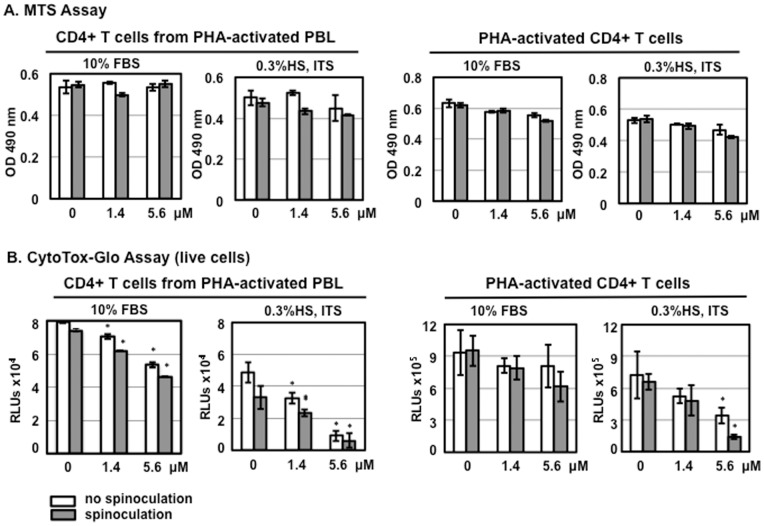
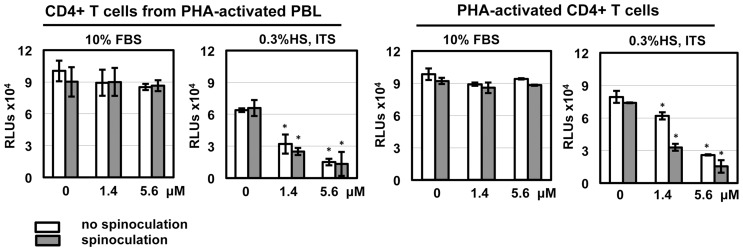
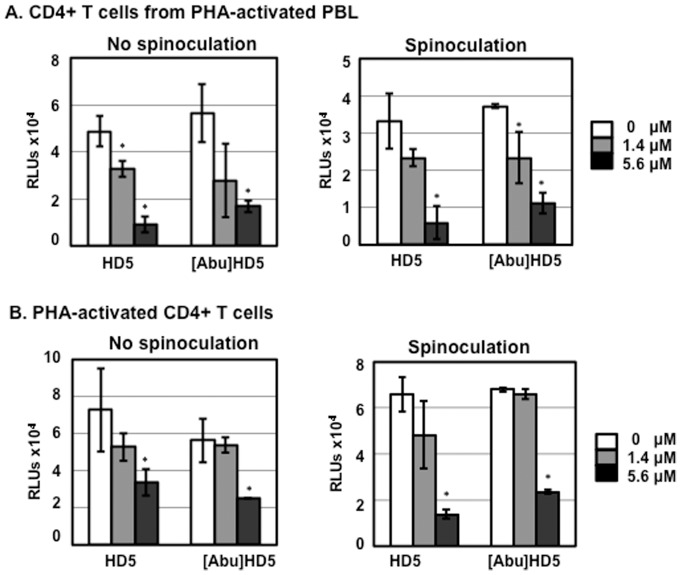
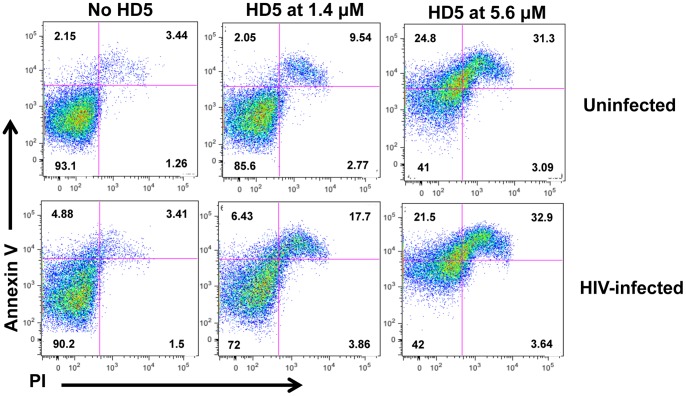
Results: Serum-deprived culture conditions were associated with anti-HIV activity. In contrast to the dependence of the HIV enhancing effect on HD5 structure, the anti-HIV activity in serum-deprived primary CD4+ T cells was independent of HD5 structure as the linear peptide [Abu] HD5 exhibited similar anti-HIV activity. Under serum deprived conditions, HD5 blocked CD4-receptor-independent HIV-1vsv infection before or after viral entry. We found that HD5 and its linear form induced significant cell death in primary CD4+ T cells under serum-deprived culture conditions. HD5-mediated apoptosis was observed as early as 2 h after addition of defensins to serum-deprived primary CD4+ T cells. In contrast to primary CD4+ T cells, HD5 did not induce cytotoxicity and promote HIV infectivity of HeLa-CD4-CCR5 cells under serum-deprived conditions.
Conclusions: These results indicate that under serum-deprived culture conditions HD5 is toxic for primary CD4+ T cells, warranting caution in data interpretation.
Conflict of interest statement
Figures
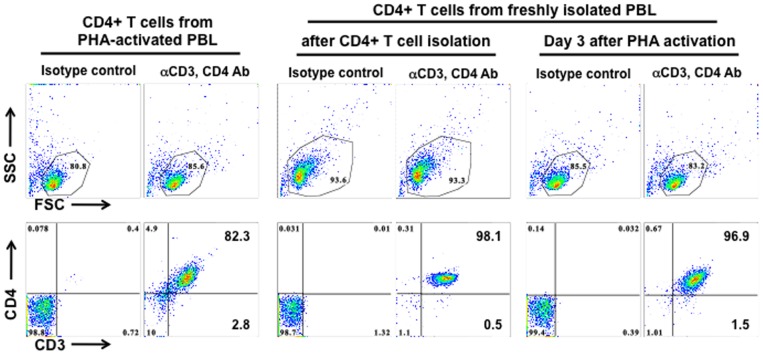
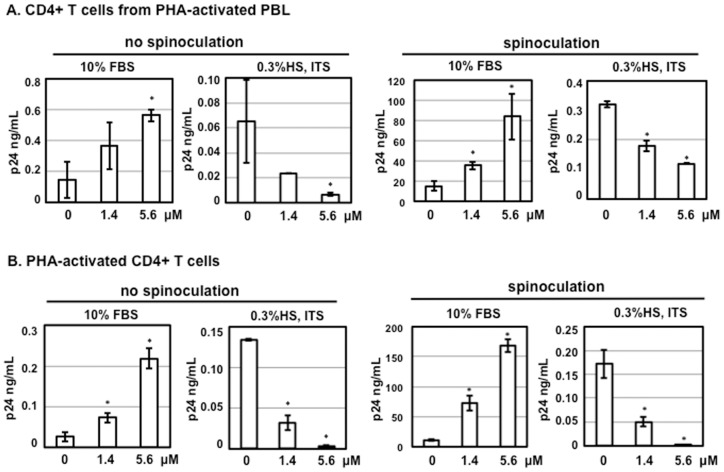
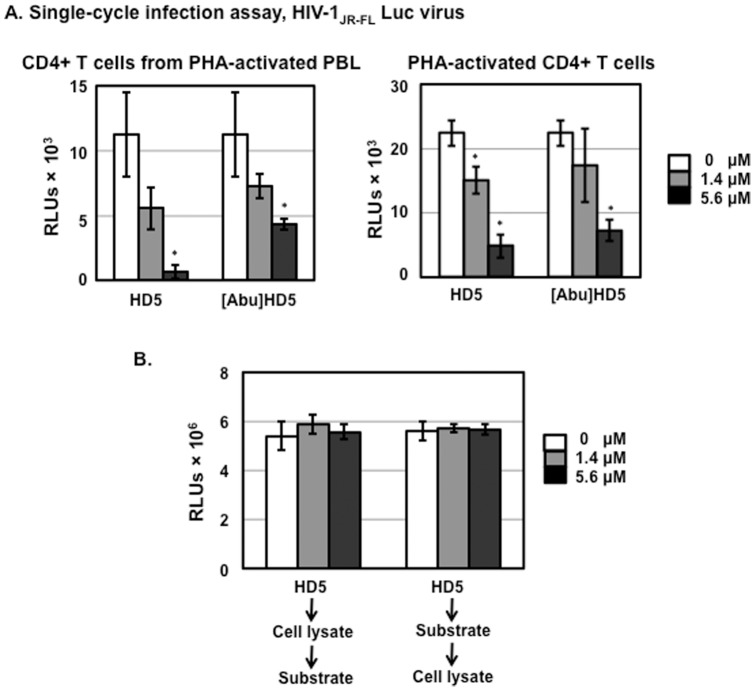
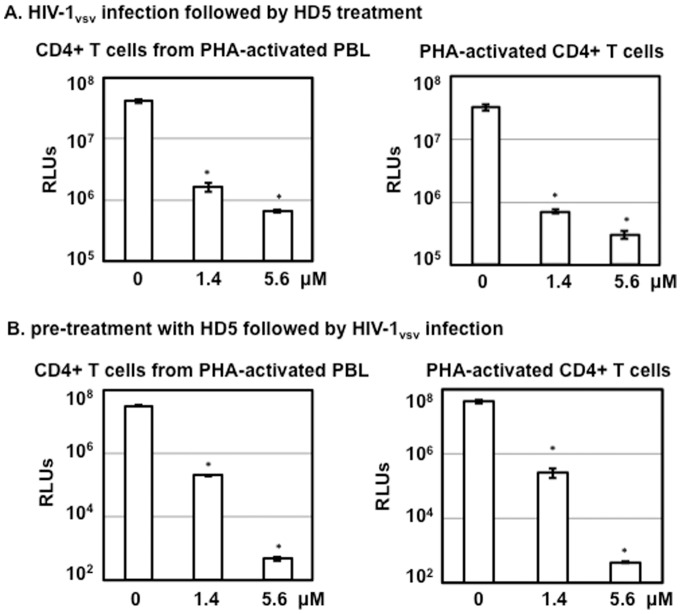
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
